Энергетический и пластический обмен
не просмотрено
Способы получения энергии живыми организмами. Типы метаболизма
Обмен веществ в организме человека
Пластический обмен
Фотосинтез и хемосинтез
Генетический код
Биосинтез нуклеиновых кислот. Репликация ДНК
Транскрипция. Созревание РНК. Типы РНК
Трансляция (биосинтез белка)
Обмен веществ. Энергетический обмен. Роль АТФ
Запись занятия: 2 часа 27 минутТеория к занятию
Обмен веществ в организме человека
Биология. Экспресс-подготовка к ЕГЭ. 11 класс.
Обмен веществ и энергии, или метаболизм — это совокупность всех химических реакций, происходящих в организме.
Обмен веществ и энергией представляет собой основу жизнедеятельности и принадлежит к критериям живого.
Нет ни одного процесса в живом организме, который бы шел без участия метаболизма, так как в основе любого физиологического процесса лежат физические и химические преобразования.
В процессе метаболизма, поступившие в организм вещества, путём химических изменений превращаются в собственные вещества тканей или в конечные продукты которые выводятся из организма. При этих химических превращениях освобождается и поглощается энергия.
Все химические реакции, проходящие в организме, являются ферментативными.
В организме осуществляется нервно-гуморальная регуляция метаболизма.
В организме динамически уравновешены пластический и энергетический обмен, входящие в состав метаболизма:
· пластический обмен = анаболизм = ассимиляция — биосинтеза органических веществ, компонентов клеток и тканей;
· энергетический обмен = катаболизм = диссимиляция — расщепление сложных молекул и компонентов клеток.

Рис. Метаболизм
Преобладание анаболических процессов обеспечивает рост, накопление массы тела, преобладание же катаболических процессов ведет к частичному разрушению тканевых структур, уменьшению массы тела. При катаболизме происходит превращение химической энергии соединений, освобождаемой при их расщеплении, в тепловую, механическую и, частично, в электрическую энергию.
ФУНКЦИИ ОБМЕНА ВЕЩЕСТВ
· Получение энергии для функционирования организма;
· Получение строительного материала для роста и восстановления организма: синтез белков, нуклеиновых кислот, углеводов, липидов и других клеточных компонентов из полученных с пищей веществ;
· Запасание питательных веществ на "черный день";
· Выведение продуктов метаболизма.
скорость обмена веществ
Скорость переноса веществ и энергии из среды в организм точно уравновешивается скоростью переноса из организма в среду. Интенсивность обмена веществ оценивают по общему расходу энергии, и она может меняться в зависимости от многих условий и в первую очередь от физической работы. Однако и в состоянии полного покоя обмен веществ и энергии не прекращается, и для обеспечения непрерывного функционирования внутренних органов, поддержания тонуса мышц и прочее расходуется некоторое количество энергии.
У молодых мужчин основной обмен веществ составляет 1300 — 1600 килокалорий (кКал) в сутки.
У женщин величина основного обмена 1100 — 1400 кКал.
ФАКТОРЫ, ВЛИЯЮЩИЕ НА СКОРОСТЬ МЕТАБОЛИЗМА
· физическая нагрузка: при интенсивной физической нагрузки расход энергии (катаболизм) по сравнению со средними энергозатратами может увеличиться в 10 раз, а в очень короткие периоды (например, плавание на короткие дистанции) даже в 100 раз;
· возраст: начиная с 5 лет величина основного обмена веществ неуклонно снижается;
· температура: с повышением температуры тела на 1 градус величина основного обмена веществ возрастает на 13%; возрастание интенсивности обмена веществ наблюдается так же при снижении температуры окружающей среды ниже зоны комфорта. Это адаптационный процесс, связанный с необходимостью поддерживать постоянную температуру тела.
незаменимые вещества
Не все вещества, необходимые для построения собственных сложных веществ, организм человека может синтезировать сам. Существуют так называемые незаменимые вещества.
1. Незаменимые аминокислоты. Из 20 аминокислот, входящих в состав белков, человек может образовывать только 12, остальные 8 должны поступать с пищей. Для людей незаменимыми аминокислотами являются лейцин, изолейцин, валин, метионин, лизин, треонин, фенилаланин, триптофан.
2. Полиненасыщенные жирные кислоты. Клетки человека не способны образовывать более одной двойной связи в жирных кислотах, поэтому они должны поступать с пищей.
3. Витамины. Эти вещества нужны в очень небольших количествах (мкг или мг). Большинство из них участвуют в образовании ферментов; при их недостатке развиваются специфические заболевания -- гиповитаминозы.

Способы получения энергии живыми организмами. Типы метаболизма
Биология. Экспресс-подготовка к ЕГЭ. 11 класс.
ТИПЫАНАБОЛИЗМА
Реакции анаболизма у разных организмов могут иметь некоторые отличия. Прежде всего, одни организмы могут образовывать все необходимые им органические вещества из неорганических веществ окружающей среды. Такие организмы называются автотрофами. Другие организмы, называемые гетеротрофами, не способны образовывать органические вещества из неорганических и должны получать их из окружающей среды.
Среди гетеротрофов есть такие, которые нуждаются лишь в одном или нескольких простых органических веществах. Например, пекарские дрожжи могут жить, получая только глюкозу и минеральные соли, а кишечная палочка — только ацетат натрия (соль уксусной кислоты).
Другие гетеротрофы не могут образовывать целый ряд необходимых им органических веществ. Например, млекопитающие нуждаются в 10 незаменимых аминокислотах, ряде ненасыщенных жирных кислот и витаминов. Некоторые внутриклеточные паразиты не могут синтезировать вообще никаких низкомолекулярных предшественников, получая их из клетки-хозяина.
ОТНОШЕНИЕ ОРГАНИЗМОВ К КИСЛОРОДУ
Энергетический обмен также различен у разных групп организмов. Гетеротрофные организмы получают энергию за счет окисления органических веществ, поступающих извне. Хорошим окислителем является молекулярный кислород, при кислородном окислении выделяется много энергии (все вы об этом знаете, т. к. видели окисление органических веществ при горении, например, в костре). Однако кислород в силу своей реакционной способности повреждает важные вещества клетки (белки, липиды, ДНК) и поэтому токсичен.
По отношению к молекулярному кислороду организмы подразделяются на:
1. Анаэробы. Они способны жить в отсутствие кислорода, осуществляя реакции внутримолекулярного окисления и восстановления или используя в качестве окислителей другие вещества среды, например нитраты (так называемое нитратное дыхание).
Анаэробов можно подразделить на:
а) облигатные анаэробы, для которых даже низкие концентрации молекулярного кислорода токсичны. Таковы, например, бактерии родов клостридий и бифидум, взрослые аскариды;
б) факультативные анаэробы могут нормально жить и размножаться как в присутствии, так и в отсутствие кислорода, например дрожжи.
2. Аэробы — большинство ныне живущих гетеротрофных организмов, которые осуществляют энергетический обмен, окисляя органические вещества молекулярным кислородом. В клетках таких организмов обязательно существуют системы защиты от окисления клеточных структур кислородом. Это, во-первых, ферменты, разрушающие активные формы кислорода (примеры таких ферментов — супероксиддисмутаза, каталаза), а во-вторых, антиоксиданты — молекулы, которые сами окисляются, «гася» свободные радикалы.
КЛАССИФИКАЦИЯ ОРГАНИЗМОВ ПО ТИПУ МЕТАБОЛИЗМА
Типы метаболизма классифицируются в зависимости от используемых источников углерода и энергии.
1. По используемому источнику С организмы делятся на автотрофы и гетеротрофы.
Автотрофы используют неорганический источник углерода (CO2), а гетеротрофы нуждаются в органических источниках углерода.
2. По используемому источнику энергии организмы делятся на фототрофы и хемотрофы.
Фототрофы используют энергию света, а хемотрофы — энергию, выделяющуюся при протекании тех или иных химических реакций.

АВТОТРОФЫ
Автотрофные организмы также используют различные способы получения энергии. Одни из них получают энергию для восстановления углекислого газа до органических соединений в виде света. Они называются фотоавтотрофы. К этой группе относятся растения и некоторые группы бактерий, прежде всего цианобактерии. Эти организмы являются основным источником органического вещества в биосфере. Понятно, что их распространение ограничено местами, в которых имеется достаточно света, т. е. поверхностью суши и верхними слоями водоемов. В биоценозах они выполняют функцию продуцентов.
Другая группа автотрофов использует для своей жизнедеятельности энергию, выделяющуюся при окислении неорганических веществ. Их называют хемоавтотрофами. Представителями этой группы являются только бактерии. Каждый вид таких бактерий способен использовать только один источник энергии. Понятно, что используемые ими вещества должны в достаточных количествах присутствовать в окружающей среде.
Хемоавтотрофные организмы классифицируются в зависимости от окисляемых веществ:
1. Организмы, окисляющие восстановленные соединения серы — сульфиды и сероводород (S2−). Бактерии, живущие за счет этих соединений, встречаются в водоемах, находящихся в местах выхода сульфидных руд и минеральных источников, содержащих сероводород. Многочисленны они в различных вулканических источниках и озерах, причем часто обитают в водоемах с высокой температурой. Эти бактерии обеспечивают в качестве единственных продуцентов существование глубоководных биоценозов «черных курильщиков».
2. Другая группа хемоавтотрофов окисляет двухвалентное железо до трехвалентного (Fe2+ -> Fe3+). Они встречаются в водах минеральных источников, а также в заболоченных водоемах, где в восстановительной среде донных отложений происходит восстановление солей железа. В древних океанах окисленные ими соединения железа выпадали в осадок и образовали многие из современных месторождений руд этого металла.
3. В почвах широко представлены бактерии, окисляющие соединения азота. Попадающие в почву остатки животных и растений при разложении выделяют азот в виде ионов аммония. Одни группы бактерий окисляют их до молекулярного азота, другие — до нитритов. Третья группа бактерий окисляет нитриты до нитратов. Эти группы хемоавтотрофных бактерий наиболее многочисленны в почвах, богатых органикой, таких как черноземы и красноземы. Они играют большую роль в формировании плодородия почв и круговороте азота в биосфере.
Фотосинтез и хемосинтез
Биология. Экспресс-подготовка к ЕГЭ. 11 класс.
ОПРЕДЕЛЕНИЕ
Фотосинтез — процесс синтеза органических веществ из неорганических (воды и углекислого газа) с использованием энергии солнечного света.
Первые опыты по фотосинтезу были проведены Джозефом Пристли в XVIII в., когда он обратил внимание на «порчу» воздуха в герметичном сосуде горящей свечой (воздух переставал быть способен поддерживать горение, помещенные в него животные задыхались) и «исправление» его растениями. Пристли сделал вывод, что растения выделяют кислород, который необходим для дыхания и горения.

ОПРЕДЕЛЕНИЕ
Фототрофы — организмы, использующие фотосинтез.
Фотоавтотрофами являются большинство растений и некоторые бактерии.
фотосинтетические пигменты
Фотосинтез может осуществляться только с помощью определенных веществ — пигментов.
Фотосинтетические пигменты высших растений делятся на две группы: хлорофиллы и каротиноиды.
Роль этих пигментов состоит в том, чтобы поглощать свет и превращать его энергию в химическую энергию. Пигменты локализованы в мембранах хлоропластов, и хлоропласты обычно располагаются в клетке так, чтобы их мембраны находились под прямым углом к источнику света, что гарантирует максимальное поглощение света.

Данные о наличии у красных водорослей хлорофилла d в настоящее время не подтверждаются — по всей видимости, в экспериментах пробы были загрязнены цианобактериями, у которых этот тип хлорофилла действительно встречается. Однако во многих источниках можно по-прежнему встретить информацию о наличии хлорофилла d у красных водорослей.
У растений в фотосинтезе участвует пигмент хлорофилл, который содержится в хлоропластах на мембранах тилакоидов. Хлорофилл придает хлоропластам и всему растению зеленую окраску.

По химическому строению хлорофилл напоминает белок крови — гемоглобин. Он имеет такое же порфириновое кольцо, только у гемоглобина в центре этого кольца находится атом железа, а у хлорофилла — магний. Порфириновое кольцо представляет собой почти плоскую пластинку, от которой отходят две органических цепочки, одна из которых очень длинная, отходит под углом, и с ее помощью хлорофилл крепится к мембранам.

Уникальное свойство хлорофилла: он умеет поглощать энергию солнечного света, переходя в возбужденное состояние.
Хлорофиллы поглощают главным образом красный и сине-фиолетовый свет. Зеленый свет они отражают и потому придают растениям характерную зеленую окраску, если только ее не маскируют другие пигменты. Существует несколько форм этого пигмента, которые различаются своим расположением в мембране. Каждая форма слегка отличается от других и по положению максимума поглощения в красной области; например, этот максимум может быть при 670, 680, 690 или 700 нм.
Хлорофилл а — единственный пигмент, который имеется у всех фотосинтезирующих растений и играет у них центральную роль в фотосинтезе.
Спектры поглощения хлорофиллов a и b и спектр каротиноидов.

Каротиноиды — пигменты желтого, красного и оранжевого цвета. Они придают окраску цветкам и плодам растений. Каротиноиды постоянно присутствуют в листьях, но незаметны из-за присутствия хлорофилла. Зато осенью, когда хлорофилл разрушается, каротиноиды становятся хорошо видны. Именно они придают листьям желтую и красную окраску.
Функции каротиноидов:
· поглощают солнечный свет (особенно в коротковолновой — сине-фиолетовой — части спектра) и поглощенную энергию передают хлорофиллу;
· защищают хлорофилл от избытка света и от окисления кислородом, выделяющимся при фотосинтезе.
ПИГМЕНТЫБАКТЕРИЙ
фотосинтез
Процесс фотосинтеза включает 2 фазы:
световая фаза:
· на свету;
· на мембранах тилакоидов;
темновая фаза:
· на свету и в темноте;
· в строме хлоропласта.

CВЕТОВАЯ ФАЗА ФОТОСИНТЕЗА
В хлоропластах содержится очень много молекул хлорофилла. Сам процесс происходит примерно в 1 % молекул хлорофилла. Другие же молекулы хлорофилла, каротиноидов и других веществ образуют особые антенные, а также светособирающие комплексы (ССК). Они, как антенны, поглощают кванты света и передают возбуждение в особые реакционные центры. Эти центры находятся в фотосистемах, которых у растений две: фотосистема II и фотосистема I. В них имеются особые молекулы хлорофилла: соответственно, в фотосистеме II — P680, а в фотосистеме I — P700. Они поглощают свет именно такой длины волны (680 и 700 нм).
· Молекулы хлорофилла двух фотосистем поглощают квант света. Один электрон каждой из них переходит на более высокий энергетический уровень (возбуждается).
· Возбужденные электроны обладает очень высокой энергией. Они отрываются и поступают в особую цепь переносчиков в мембранах тилакоидов — молекулы НАДФ+, превращая их в восстановленный НАДФ. Таким образом, энергия света превращается в энергию восстановленного переносчика.
· В молекулах хлорофилла на месте электронов после их отрыва образуются "дырки" с положительным зарядом.
· Фотосистема I восполняет потерю электронов через систему переносчиков электронов от фотосистемы II.
· Фотосистема II забирает электрон у воды (фотолиз воды), при этом образуются ионы водорода.
· Фотолиз воды — процесс распада воды под действием солнечного света.

· Побочным продуктом распада воды является кислород, выделяющийся в атмосферу.
· НН+, образовавшиеся при фотолизе воды, переносятся в полость тилакоида.
· В полости тилакоида накапливается большой избыток ионов водорода, что приводит к созданию на мембране тилакоида крутого градиента концентрации этих ионов.
· Он используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфата.
· Происходит перенос ионов водорода НН+ через мембрану восстановленным переносчиком НАДФ (никотинамидадениндинуклеотидфосфатом) с образованием НАДФ*Н.
Таким образом, энергия света запасается в световой фазе фотосинтеза в виде двух типов молекул: восстановленного переносчика НАДФ*Н и макроэргического соединения АТФ. Кислород, выделяющийся при этом, является с точки зрения фотосинтеза побочным продуктом.
Роль световой фазы:
· перенос протонов водорода через систему переносчиков с образованием энергии АТФ;
· образование НАДФ*Н;
· выделение молекулярного кислорода в атмосферу.
ТЕМНОВАЯ ФАЗА ФОТОСИНТЕЗА
Для темновой фазы фотосинтеза обязательными компонентами являются АТФ и НАДФ*Н (из световой фазы), углекислый газ (из атмосферы) и вода. Происходит в строме хлоропласта.
В темновой фазе с участием АТФ и НАДФ*Н происходит восстановление CO2 до глюкозы (C6H12O6).
Хотя свет не требуется для осуществления данного процесса, он участвует в его регуляции.
Растение постоянно поглощает углекислый газ из атмосферы. Для этой цели на поверхности листа имеются специальные структуры — устьица. Когда они открываются, CO2 поступает внутрь листа, растворяется в воде и восстанавливается до глюкозы с помощью НАДФ и АТФ.
Избыток глюкозы запасается в виде крахмала. Именно в виде этих органических веществ растение накапливает энергию. Только небольшая их часть остается в листе и используется для его нужд. Остальные же углеводы путешествуют по ситовидным трубкам флоэмы по всему растению и поступают именно туда, где больше всего нужна энергия, например в точки роста.
ЦИКЛ КАЛЬВИНА
С3- И С4-ФОТОСИНТЕЗ
Суммарное уравнение фотосинтеза выглядит следующим образом:
6СО2 + 6Н2О+ энергия света → С6Н12О6 + 6О2.
ЗНАЧЕНИЕ ФОТОСИНТЕЗА
· Фотосинтез является основным источником органического вещества на Земле, то есть обеспечивает живые организмы веществом и энергией.
· Он служит источником кислорода, составляющего 20 % атмосферы Земли. Весь атмосферный кислород образовался в результате фотосинтеза. До появления организмов, осуществляющих фотосинтез с выделением кислорода (около 3 млрд лет назад), атмосфера Земли не содержала этого газа.
хемосинтез
ОПРЕДЕЛЕНИЕ
Хемосинтез — способ автотрофного питания, при котором источником энергии для синтеза органических веществ служит окисление неорганических соединений.
К хемосинтетикам (хемотрофам) относятся только некоторые бактерии и археи.
Явление хемосинтеза было открыто в 1887 г. русским ученым С. Н. Виноградским.
Процесс хемосинтеза, при котором из CO2 образуется органическое вещество, протекает аналогично темновой фазе фотосинтеза, только используется АТФ, полученный не из солнечной энергии, а из энергии химических связей неорганического вещества (при окислении серы, железа, аммиака и т.п.).
Благодаря жизнедеятельности бактерий-хемосинтетиков в природе накапливаются большие запасы селитры и болотной руды.
ХЕМОСИНТЕЗИРУЮЩИЕ БАКТЕРИИ
· Нитрифицирующие бактерии получают энергию для синтеза органических веществ, окисляя аммиак до азотистой, а затем до азотной кислоты:
2NH3 + 3O2 → 2HNO2 + 2H20 + Q;
2HNO2 + O2 → 2HNO3 + Q.
· Серобактерии получают энергию, окисляя сероводород до сульфатов:
2H2S + 02 → 2H20 + 2S + Q;
S+ 3O2 + 2H2O → 2H2SO4 + Q.
· Водородные бактерии получают энергию, окисляя водород до воды:
2НН2 + O2 → 2H2O + Q.
· Железобактерии получают энергию, окисляя Fe2+ до Fe3+:
4Fe(HCO3)2+6H_{2}O$ + 02 → 4Fe(OH)3 + 4H2CO3 +4CO2 + Q.
При этой реакции энергии выделяется немного, поэтому железобактерии окисляют большое количество закисного железа.
Полученная в реакциях окисления неорганических соединений энергия переводится в энергию макроэнергетических связей АТФ.
РОЛЬ ХЕМОСИНТЕТИКОВ
· участвуют в круговороте серы, азота, железа и др.;
· уничтожают в природе ядовитые вещества: аммиак и сероводород;
· нитрифицирующие бактерии превращают аммиак в нитриты и нитраты, усваиваемые растениями;
· серобактерии используются для очистки сточных вод.
Хемоорганогетеротрофы
Хемоорганогетеротрофы — организмы, использующие для синтеза собственных органических веществ энергию, получаемую при окислении органических веществ пищи в процессе дыхания. К хемоорганогетеротрофам относятся животные, грибы и некоторые бактерии (например, клубеньковые азотфиксирующие бактерии).
$
Генетический код
Биология. Экспресс-подготовка к ЕГЭ. 11 класс.
Веществами, ответственными за хранение и передачу генетической информации, являются нуклеиновые кислоты (ДНК и РНК).
Все функции клеток и организма в целом определяются набором белков, обеспечивающих
· образование клеточных структур,
· синтез всех других веществ (углеводов, жиров, нуклеиновых кислот),
· протекание процессов жизнедеятельности.
В геноме содержится информация о последовательности аминокислот во всех белках организма. Именно эта информация и называется генетической информацией.
За счёт регуляции генов регулируется время синтеза белков, их количество, место нахождения в клетке или в организме в целом. Во многом за это отвечают регуляторные участки ДНК, усиливающие и ослабляющие экспрессию генов в ответ на те или иные сигналы.
Информация о белке может быть записана в нуклеиновой кислоте только одним способом — в виде последовательности нуклеотидов. ДНК построена из 4 видов нуклеотидов (А, Т, Г, Ц), а белки — из 20 видов аминокислот. Таким образом, возникает проблема перевода четырёхбуквенной записи информации в ДНК в двадцатибуквенную запись белков. Соотношения, на основе которых осуществляется такой перевод, называются генетическим кодом.
Первым проблему генетического кода теоретически рассмотрел выдающийся физик Георгий Гамов. Генетический код обладает определённым набором свойств, которые будут рассмотрены ниже.
ПОЧЕМУ НЕОБХОДИМ ГЕНЕТИЧЕСКИЙ КОД?
СВОЙСТВА ГЕНЕТИЧЕСКОГО КОДА
КОЛЛИНЕАРНОСТЬ (ЛИНЕЙНОСТЬ)
Сначала мы рассмотрим, как в последовательности нуклеотидов записана последовательность аминокислот в белках. Логично предположить, что, поскольку последовательности нуклеотидов и аминокислот линейны, то между ними существует линейное соответствие, т. е. расположенным рядом нуклеотидам в ДНК соответствуют расположенные рядом аминокислоты в полипептиде. На это же указывает линейный характер генетических карт. Доказательством такого линейного соответствия, или коллинеарности, является совпадение линейного расположения мутаций на генетической карте и аминокислотных замен в белках мутантных организмов.
ТРИПЛЕТНОСТЬ
При рассмотрении свойств кода реже всего встает вопрос о кодовом числе. Необходимо закодировать 20 аминокислот четырьмя нуклеотидами. Очевидно, что 1 нуклеотид не может кодировать 1 аминокислоту, т. к. тогда было бы возможно закодировать только 4 аминокислоты. Для того чтобы закодировать 20 аминокислот, нужны комбинации из нескольких нуклеотидов. Если взять комбинации из двух нуклеотидов, то мы получим 16 различных комбинаций (42 = 16). Этого недостаточно. Комбинаций из трёх нуклеотидов будет уже 64 (43 = 64), т. е. даже больше, чем нужно. Понятно, что комбинации из большего числа нуклеотидов тоже могли бы быть использованы, но из соображений простоты и экономии они маловероятны, т. е. код триплетный.
ВЫРОЖДЕННОСТЬ И ОДНОЗНАЧНОСТЬ
В случае 64 комбинаций возникает вопрос, все ли комбинации кодируют аминокислоты или каждой аминокислоте соответствует только один триплет нуклеотидов. Во втором случае большая часть триплетов была бы бессмысленной, а замены нуклеотидов в результате мутаций в двух третях случаев приводили бы к потере белка. Это не соответствовует наблюдаемым частотам потери белка при мутациях, что указывает на использование всех или почти всех триплетов. В дальнейшем было выяснено, что существуют три триплета, не кодирующие аминокислот. Они служат для того, чтобы обозначать конец полипептидной цепочки. Их называют стоп-кодонами. 61 триплет кодирует различные аминокислоты, т. е. одна аминокислота может кодироваться несколькими триплетами. Это свойство генетического кода называется вырожденностью. Вырожденность имеет место только в направлении от аминокислот к нуклеотидам, в обратном направлении код однозначен, т.е. каждый триплет кодирует одну определённую аминокислоту.
ЗНАКИ ПРЕПИНАНИЯ
Важный вопрос, решить который теоретически оказалось невозможным, каким образом триплеты, кодирующие соседние аминокислоты, отделяются друг от друга, т. е. есть ли в генетическом тексте знаки препинания.
ОТСУТСТВИЕ ЗАПЯТЫХ — ЭКСПЕРИМЕНТЫ
НЕПЕРЕКРЫВАЕМОСТЬ
Гамов предположил, что код перекрывающийся, т. е. первый, второй и третий нуклеотиды кодируют первую аминокислоту, второй, третий и четвёртый — вторую аминокислоту, третий, четвёртый и пятый — третью и т. д. Такая гипотеза создавала видимость решения пространственных затруднений, но создавала другую проблему. При таком кодировании за данной аминокислотой не могла идти любая другая, так как в кодирующем её триплете два первых нуклеотида уже были определены, и число возможных триплетов снижалось до четырёх. Анализ последовательностей аминокислот в белках показал, что встречаются все возможные пары соседних аминокислот, т. е. код должен быть неперекрывающимся.
УНИВЕРСАЛЬНОСТЬ
Еще одним свойством генетического кода, постулированным из теоретических соображений, является его универсальность. Предполагалось, что все виды живых организмов произошли в результате эволюции от одного общего предка, поэтому они имеют одинаковый генетический код. Это положение подтвердилось дальнейшими исследованиями. Оно имеет большое практическое значение, т. к. благодаря универсальности генетического кода мы можем заставить гены одного организма работать в другом организме и производить функционально активные белки. Это позволяет с помощью методов генетической инженерии получать в бактериях белки человека, нужные для медицинских целей, например, инсулин или гормон роста. Однако универсальность генетического кода оказалась не абсолютной. Известно несколько генетических систем, в которых генетический код немного отличается от универсального. Прежде всего это митохондрии. Кроме того, отклонения от универсального генетического кода найдены у некоторых инфузорий и паразитических бактерий. Однако во всех этих случаях отклонения незначительны и, очевидно, возникли вторично на основе универсального кода.
РАСШИФРОВКА КОДА
Когда основные свойства генетического кода были изучены, начались работы по его расшифровке и были определены значения всех триплетов (см. рис.). Триплет, кодирующий определённую аминокислоту, получил название кодона. Как правило, указываются кодоны в мРНК, иногда — в смысловой цепи ДНК (те же кодоны, но с заменой У на Т). Для некоторых аминокислот, например, метионина, существует только один кодон. Другие имеют по два кодона (фенилаланин, тирозин). Есть аминокислоты, которые кодируются тремя, четырьмя и даже шестью кодонами. Кодоны одной аминокислоты похожи друг на друга и, как правило, отличаются одним последним нуклеотидом. Это делает генетический код более устойчивым, так как замена последнего нуклеотида в кодоне при мутациях не ведёт к замене аминокислоты в белке. Знание генетического кода позволяет нам, зная последовательность нуклеотидов в гене, выводить последовательность аминокислот в белке, что широко используется в современных исследованиях.

Биосинтез нуклеиновых кислот. Репликация ДНК
Биология. Экспресс-подготовка к ЕГЭ. 11 класс.
БИОСИНТЕЗ НУКЛЕИНОВЫХ КИСЛОТ. СИНТЕЗ ДНК
Принцип комплементарности лежит в основе процессов синтеза всех нуклеиновых кислот. Впервые модель синтеза нуклеиновой кислоты предложили Дж. Уотсон и Ф. Крик одновременно с моделью двойной спирали ДНК. Они предположили, что благодаря принципу комплементарности старая молекула может служить матрицей для синтеза новых идентичных молекул. Это возможно благодаря тому, что водородные связи между цепями ДНК гораздо слабее ковалентных и могут быть разрушены. На это тратится энергия — в клетке в качестве ее источника используется АТФ, в пробирке цепи можно разделить простым нагреванием. Этот процесс называется плавлением, или денатурацией ДНК.

После разделения цепей «обнажаются» азотистые основания, не связанные более водородными связями. Они могут провзаимодействовать по принципу комплементарности с новыми нуклеотидами, которые станут звеньями дочерних цепей. При этом каждая из двух материнских цепочек становится матрицей для синтеза дочерней цепи. В результате получаются две дочерние двойные спирали ДНК, идентичные исходной молекуле. Принцип матричного синтеза лежит в основе синтеза всех существующих в клетке нуклеиновых кислот (как ДНК, так и РНК).
В дальнейшем оказалось, что полного расхождения нитей ДНК не происходит, расплетается только небольшой фрагмент ДНК, на нем происходит поочередное присоединение нуклеотидов. Расплетенный участок материнской молекулы, где идет наращивание новых цепей, называют репликативной вилкой. Каждый очередной новый нуклеотид подбирается по принципу комплементарности к находящемуся против него нуклеотиду матричной нити (см. рис.). Для синтеза ДНК используются нуклеотиды, содержащие дезоксирибозу, а в качестве азотистых оснований аденин, гуанин, цитозин и тимин. Этот процесс называется удвоением, или репликацией ДНК. Его осуществляет фермент ДНК-зависимая ДНК-полимераза (ДНК-зависимая означает, что она использует ДНК в качестве матрицы), или просто ДНК-полимераза (но существуют и РНК-зависимые ДНК-полимеразы, например у ретровирусов).
направление синтеза
Синтез новой цепи начинается с 5’-конца, новые нуклеотиды присоединяются всегда к 3’-концевому нуклеотиду, к его свободной ОН-группе. При этом комплекс белков, синтезирующий ДНК, двигается вдоль матричной нити в направлении от 3' к 5', постепенно расплетая новые участки двойной спирали. Новые данные экспериментов показывают, что, скорее, наоборот, ДНК протягивается сквозь статичную «фабрику репликации» — совокупность осуществляющих репликацию белков. Так продолжается до полного завершения удвоения всей молекулы ДНК.
ферменты репликативной вилки
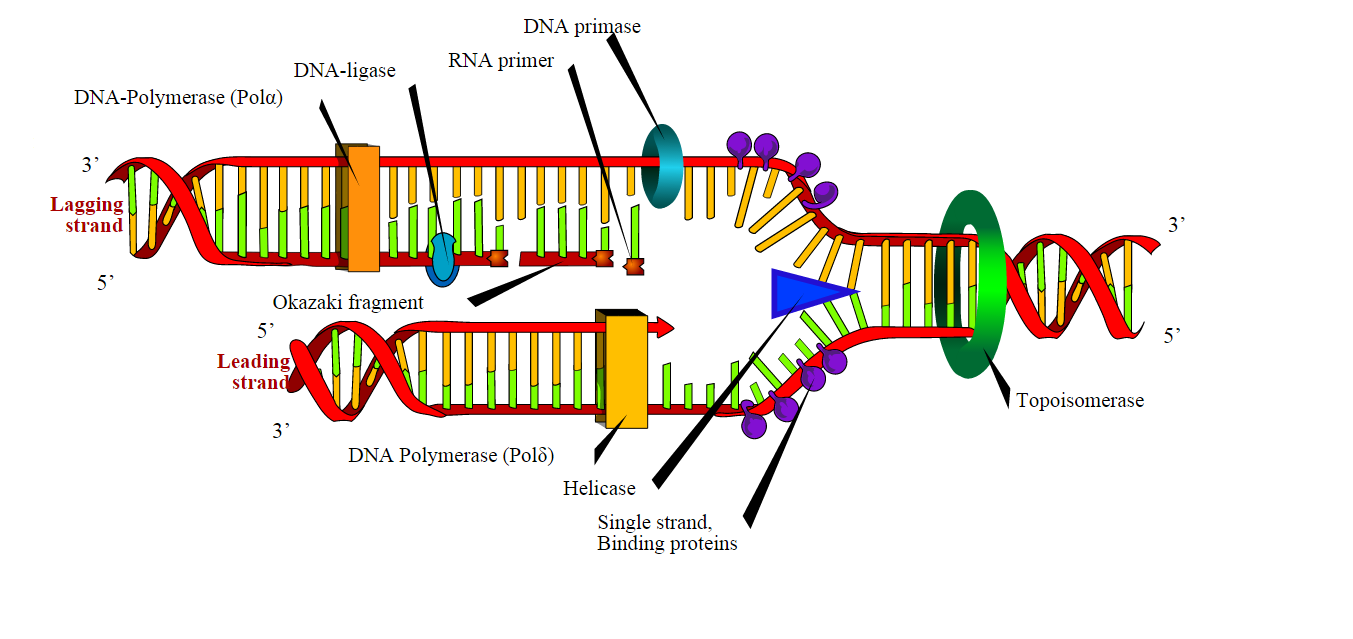
Расплетание материнской молекулы АТФ-зависимо осуществляет фермент ДНК-хеликаза (от helix — спираль). На обнаженных матричных цепях праймаза синтезирует РНК-затравки, или праймеры — короткие участки РНК. 3'-концы праймеров удлиняются ДНК-полимеразами. У прокариот основной репликативной полимеразой является РНК-полимераза III. У эукариот больше типов ДНК-полимераз, они обозначаются греческими буквами.
Основная репликативная ДНК-полимераза должна быть процессивна, то есть синтезировать очень длинный фрагмент ДНК без «соскоков» с матрицы и обладать свойством проверки ошибок — proofreading. Оно заключается в том, что при случайном присоединении некомплементарного нуклеотида ДНК-полимераза отщепляет его от растущей цепи. Это обеспечивает fidelity — точность синтеза.
Особенностью синтеза ДНК является невозможность его начала «с нуля». Все ДНК-полимеразы узнают и удлиняют уже существующий 3'-конец молекулы, спаренной с матрицей. Поэтому им нужна затравка, или праймер — любая молекула, которая комплементарно связана с матрицей и 3'-конец которой можно удлинять, руководствуясь этой матрицей. Так как синтез РНК происходит беззатравочно, в клетке роль праймеров играет РНК. Эти короткие фрагменты РНК синтезирует специфическая РНК-полимераза — праймаза.
На одной из цепей направление синтеза ДНК (от 3' к 5' по матрице) совпадает с направлением расплетания ДНК (движения вилки). Эта цепь называется лидирующей, на ней синтез происходит непрерывно. На другой цепи направление синтеза ДНК противоположно направлению расплетания, так что синтез происходит фрагментами, по мере расплетания все новых и новых участков матрицы. Эта цепь называется отстающей, а фрагменты, по имени открывшего их исследователя, называют фрагментами Оказаки. Каждый фрагмент Оказаки начинается со своей затравки; между ними остаются незамкнутые связи (бреши). Затравки удаляются РНКазой Н (от hybrid — РНК-ДНК-гибрид), вместо нее ДНК-полимераза I (репаративная) синтезирует РНК. Фрагменты соединяются между собой ДНК-лигазой. После удаления всех затравок и залечивания всех брешей новая ДНК полностью готова.
субстраты для синтеза и энергия
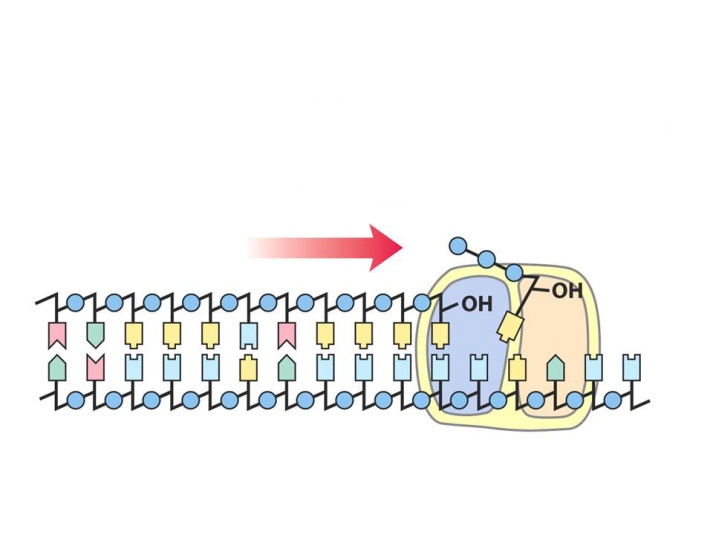
Для образования связи между нуклеотидами необходима энергия. Она поставляется в реакцию каждым приходящим нуклеотидом, т. к. используются нуклеозидтрифосфаты, имеющие макроэргические связи.
При этом от молекулы нуклеотида отщепляется два остатка фосфорной кислоты, а третий присоединяется к 3-му положению предыдущего нуклеотида (см. рис.). Синтез новой цепи начинается с 5’ конца, новые нуклеотиды присоединяются к 3’-концевому нуклеотиду. При этом комплекс белков, синтезирующий ДНК, двигается вдоль матричной нити, постепенно расплетая новые участки двойной спирали. Так продолжается до полного завершения синтеза всей молекулы ДНК. Этот процесс называется репликацией ДНК.

Синтез ДНК не является стопроцентно точным. С вероятностью порядка 10−6 в ходе синтеза происходят ошибки, в результате чего вставляется неправильный (некомплементарный) нуклеотид. В результате возникает предмутационное состояние — некомплементарная пара нуклеотидов в полученной молекуле ДНК. В клетке существует специальная система исправления ошибок в ДНК — система репарации (от англ. to repair — восстанавливать). Если система репарации исправит ошибку, то ДНК останется точной копией материнской молекулы. Если же исправление ошибки не произойдет до следующего удвоения ДНК, то изменение будет закреплено и унаследовано потомками данн