Углеводы или сахариды - одна из основных групп органических соединений организма. Они представляют собой первичные продукты фотосинтеза и исходные продукты биосинтеза других веществ в растениях (органические кислоты, аминокислоты), а также содержатся в клетках всех других живых организмов. В животной клетке содержание углеводов колеблется в пределах 1-2%, в растительной оно может достигать в некоторых случаях 85-90% массы сухого вещества.
Углеводы состоят из углерода, водорода и кислорода, причем у большинства углеводов водород и кислород содержатся в том же соотношении, что и в воде (отсюда их название - углеводы). Таковы, например, глюкоза С6Н12О6 или сахароза С12Н22О11. В состав производных углеводов могут входить и другие элементы. Все углеводы делятся на простые (моносахариды) и сложные (полисахариды).
Среди моносахаридов по числу углеродных атомов различают триозы (3С), тетрозы (4С), пентозы (5С), гексозы (6С) и гептозы (7С). Моносахариды с пятью и более атомами углерода, растворяясь в воде, могут приобретать кольцевую структуру. В природе наиболее часто встречаются пентозы (рибоза, дезоксирибоза, рибулоза) и гексозы (глюкоза, фруктоза, галактоза). Рибоза и дезоксирибоза играют важную роль в качестве составных частей нуклеиновых кислот и АТФ. Глюкоза в клетке служит универсальным источником энергии. С превращением моносахаридов связаны не только обеспечение клетки энергией, но и биосинтез многих других органических веществ, а также обезвреживание и выведение из организма ядовитых веществ, проникающих извне или образующихся в процессе обмена веществ, например, при распаде белков.
Ди- и полисахариды образуются путем соединения двух и более моносахаридов, таких, как глюкоза галактоза маноза, арабиноза или ксилоза. Так, соединяясь между собой с выделением молекулы воды, две молекулы моносахаридов образуют молекулу дисахарида. Типичными представителями этой группы веществ являются сахароза (тростниковый сахар), мальтаза (солодовый сахар), лактоза (молочный сахар). Дисахариды по своим свойствам близки к моносахаридам. Например, и те, и другие хорошо растворимы в воде и имеют сладкий вкус. К числу полисахаридов принадлежит крахмал, гликоген, целлюлоза, хитин, каллоза и др.
Основная роль углеводов связана с их энергетической функцией. При их ферментативном расщеплении и окислении выделяется энергия, которая используется клеткой. Полисахариды играют главным образом роль запасных продуктов и легко мобилизуемых источников энергии (например, крахмал и гликоген), а также используются в качестве строительного материала (целлюлоза, хитин). Полисахариды удобны в качестве запасных веществ по ряду причин: будучи нерастворимы в воде, они не оказывают на клетку ни осмотического, ни химического влияния, что весьма важно при длительном хранении их в живой клетке: твердое, обезвоженное состояние полисахаридов увеличивает полезную массу продуктов запаса за счет экономии их объема. При этом существенно уменьшается вероятность потребления этих продуктов болезнетворными бактериями и другими микроорганизмами, которые, как известно, не могут заглатывать пищу, а всасывают вещества всей поверхностью тела. И наконец, при необходимости запасные полисахариды легко могут быть превращены в простые сахара путем гидролиза.
Классификация:Моносахариды: глюкоза,фруктоза,галактоза ,Дисахариды:сахароза,лактоза.мальтоза, Полисахариды:крахмал.гликоген,пектиновыевещества,клетчатка.
2 ….. биологическое значение углеводов:
1 Углеводы выполняют пластическую функцию, то есть участвуют в построении костей, клеток, ферментов. Они составляют 2-3 % от веса.
2 Углеводы являются основным энергетическим материалом. При окислении 1 грамма углеводов выделяются 4,1 ккал энергии и 0,4 г воды.
3В крови содержится 100-110 мг глюкозы. От концентрации глюкозы зависит осмотическое давление крови.
4Пентозы (рибоза и дезоксирибоза) участвуют в построении АТФ.
5 Углеводы выполняют защитную роль в растениях.
3…….. Всасывание углеводов пищи в желудочно-кишечном тракте - это транспорт углеводов из полости желудочно-кишечного тракта во внеклеточную (межклеточную жидкость, кровь и лимфу) и внутриклеточную жидкости организма. Всасывание углеводов завершает метаболизм углеводов в системе пищеварения.
Метаболизм углеводов - это совокупность ферментативных реакций, протекающих в организме, исходным метаболитом (объектом изменения) которых являются углеводы.
Начальным этапом метаболизма углеводов является метаболизм углеводов в системе пищеварения. Это совокупность процессов переваривания полисахаридов и олигосахаридов пищевых продуктов, поступающих в пищеварительный тракт с пищей, до моносахаридов и их последующее всасывание из полости пищеварительного тракта в кровь и лимфу. Метаболизм углеводов в системе пищеварения сопряжён с метаболизмом углеводов в тканях, клетках и субклеточных структурах (митохондрии, цитозоль, эндоплазматический ретикулум и др).
Переваривание углеводов пищи в желудочно-кишечном тракте - это часть метаболизма углеводов, совокупность управляемых процессов, осуществляемых в системе пищеварения, которые представляют собой химическую переработку углеводов, поступающих в организм с пищей, для последующего их всасывания в кровь и в лимфу.
Пищевые вещества содержат крупномолекулярные белки, углеводы и липиды, которые не способны к всасыванию в кровь и лимфу из-за больших размеров своих молекул. Химическая переработка пищи в желудочно-кишечном тракте представляет собой последовательное ступенчатое ферментативное гидролитическое расщепление крупномолекулярных белков, углеводов и липидов до простых веществ, способных к всасыванию.
В числе углеводов потребляемой пищи содержатся полисахариды крахмал и гликоген. Расщепление этих углеводов начинается в полости рта и продолжается в желудке. Катализатором гидролиза является фермент α‑амилаза слюны. При расщеплении из крахмала и гликогена образуются декстрины и в небольшом количестве - мальтоза. Пережеванная и смешанная со слюной пища проглатывается и попадает в желудок. Проглоченные пищевые массы со стороны поверхности полости желудка постепенно перемешиваются с желудочным соком, содержащим соляную кислоту. Содержимое желудка с периферии приобретает значительную кислотность (pH = 1,5 ÷ 2,5). Такая кислотность дезактивирует амилазу слюны. В то же время в толще массы желудочного содержимого амилаза слюны некоторое время продолжает действовать и происходит расщепление полисахаридов с образованием декстринов и мальтозы. Желудочный сок не содержит ферментов, расщепляющих сложные углеводы. Поэтому гидролиз углеводов с увеличением кислотности в желудке прерывается и возобновляется в двенадцатиперстной кишке.
Для расщепления каждой группы веществ существуют свои определенные ферменты: для белков – протеазы, для жиров – липазы, для углеводов – карбогидразы. Основными железами, которые вырабатывают пищеварительные ферменты, являются железы ротовой полости (слюнные железы), железы желудка и тонкого кишечника, поджелудочная железа и печень. Главную роль в этом играет поджелудочная железа, которая вырабатывает не только пищеварительные ферменты, но и гормоны, такие, как инсулин и глюкагон, участвующие в регуляции белкового, углеводного и липидного обменов.
Клеток, вырабатывающих пищеварительные ферменты в поджелудочной железе, достаточно много. Они образуют специальные скопления, от которых отходят маленькие выводные протоки; по ним передвигается секретируемый сок поджелудочной железы, представляющий собой своеобразный коктейль из разных ферментов.
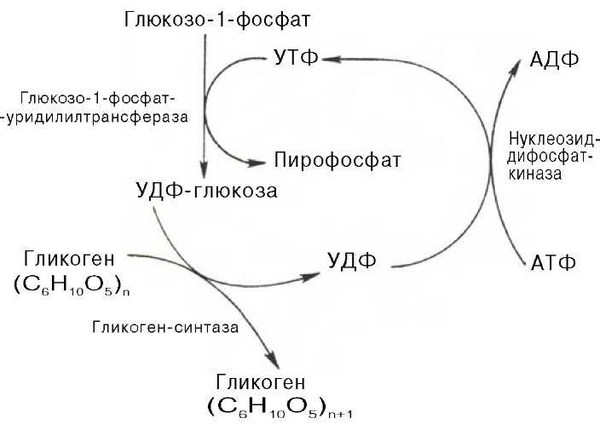
55555 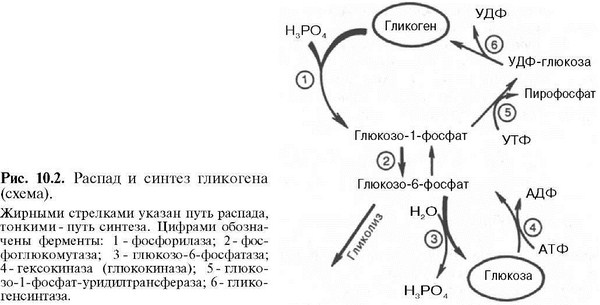
7777.. Поскольку одним из главных видов биологического «топлива» служат углеводы, мы рассмотрим биологическое окисление на примере того, как окисляется в клетках глюкоза и как запасается при этом энергия.
Окисление глюкозы сопровождается ее расщеплением, и поэтому такой процесс принято называть гликолизом (от греч. «гликис» - сладкий и «лизис» - расщепление). Анаэробный гликолиз - это неполное окисление глюкозы без участия кислорода, когда акцептором электронов служит НАД+, тогда как полный аэробный гликолиз происходит при участии кислорода.
На первом этапе гликолиза шестиуглеродная молекула глюкозы - С6Н12О6 расщепляется на две трехуглеродные молекулы пировиноградной кислоты (ПВК) - С3Н4О3. При этом происходит лишь частичное окисление глюкозы с потерей четырех атомов водорода (сравните: две молекулы ПВК С3Н4О3 * 2 = С6Н8О6 и глюкозу - С6Н12О6). За счет электронов и протонов водорода восстанавливаются две молекулы НАД+,енесенных с высоких энергетических уровней глюкозы на более низкий уровень молекул НАД+, происходит образование двух молекул АТФ из АДФ и фосфорной кислоты.
Если кислород в клетке отсутствует или его недостаточно, то две молекулы ПВК, образовавшиеся из глюкозы, восстанавливаются за счет двух НАД*Н до молочной кислоты: 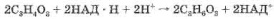
Этим завершается анаэробный гликолиз. В результате бескислородного неполного окисления глюкозы образуются всего только две молекулы АТФ.
Если в клетке уже имеется или же в нее начинает поступать кислород, то ПВК не восстанавливается до молочной кислоты, а переносится в митохондрии, где подвергается при участии кислорода полному окислению до СО2 и Н2О (как при горении). Энергетический выигрыш аэробного гликолиза, выраженный в молекулах АТФ, значительно выше, чем при анаэробном гликолизе.
Центральным звеном Г. является гликолитическая оксидоредукция (реакции 8—10), представляющаяокислительно-восстановительный процесс, протекающий с окислением 3-фосфоглицеринового альдегида до 3-фосфоглицериновой кислоты и восстановлением кофермента НАД. Эти превращения осуществляет дегидрогеназа 3-фосфоглицеринового альдегида (ДФГА) при участии фосфоглицераткиназы.
Субстратное фосфорилирование синтез богатых энергией фосфорных соединений за счёт энергии окислительно-восстановительных реакций Гликолиза (катализируемых фосфоглицеральдегиддегидрогеназой и енолазой) и при окислении α-кетоглутаровой кислоты в Трикарбоновых кислот цикле
8888888.. Регуляция гликолиза в целом, сразу для всего организма, происходит под действием гормонов, которые, влияя через молекулы вторичных посредников, изменяют внутриклеточный метаболизм.
Важное значение в стимуляции гликолиза принадлежит инсулину. Глюкагон и адреналин являются наиболее значимыми гормональными ингибиторами гликолиза.
Инсулин стимулирует гликолиз через:
1активацию гексокиназной реакции;
2стимуляцию фосфофруктокиназы;
3стимуляцию пируваткиназы.
Также на гликолиз влияют и другие гормоны. Например, соматотропин ингибирует ферменты гликолиза, а тиреоидные гормоны являются стимуляторами.
Регуляция гликолиза осуществляется через несколько ключевых этапов. Реакции, катализируемые гексокиназой (1), фосфофруктокиназой (3) и пируваткиназой (10) отличаются существенным уменьшением свободной энергии и являются практически необратимыми, что позволяет им быть эффективными точками регуляции гликолиза.
У некоторых видов микроорганизмов, живущих без доступа кислорода, имеет место только анаэробный гликолиз (например, у метанобразующих бактерий, которые живут глубоко в иле болот или озер). Мы можем короткое время обходиться без кислорода, и тогда клетки получают энергию за счет анаэробного гликолиза. Быстрый бег даже на короткие дистанции может вызвать у нас одышку. Это молочная кислота, которая образовалась при недостатке кислорода, возбуждает дыхательный центр и заставляет нас часто дышать, чтобы насытить кровь кислородом, окислить молочную кислоту, восстановить аэробный гликолиз.
99999…Аэробный распад глюкозы
Энергетическое значение аэробного распада глюкозы.
В аэробном гликолизе образуется 10 моль АТФР на 1 моль глюкозы. Так, в реакциях 7, 10 образуется 4 моль АТФ путем субстратного фосфорилирования, а в реакции 6 синтезируется 6 моль АТФ (на 2 моль глицероальдегидфосфата) путем окислительного фосфорилирования.
Баланс аэробного гликолиза.
Суммарный эффект аэробного гликолиза составляет 8 моль АТФ, так как в реакциях 1 и 3 используется 2 моль АТФ. Дальнейшее окисление двух моль пируват в общих путях катаболизма сопровождается синтезом 30 моль АТФ (по 15 моль на каждую молекулу пирувата. Следовательно, суммарный энергетический эффект аэробного распада глюкозы до конечных продуктов составляет 38 моль АТФ.
Аэробный распад глюкозы можно выразить суммарным уравнением:
С6Н12О6 + 6 О2 → 6 СО2 + Н2О + 2820 кДж/моль.
Этот процесс включает несколько стадий (рис. 7-33).
1.Аэробный гликолиз - процесс окисления глюкозы с образованием двух молекул пирувата;
2.Общий путь катаболизма, включающий превращение пирувата в ацетил-КоА и его дальнейшее окисление вцитратом цикле;
3.ЦПЭ на кислород, сопряжённая с реакциями дегидрирования, происходящими в процессе распада глюкозы.
Аэробным гликолизом называют процесс окисления глюкозы до пировиноградной кислоты, протекающий в присутствии кислорода. Все ферменты, катализирующие реакции этого процесса, локализованы в цитозоле клетки.
1. Этапы аэробного гликолиза
Подготовительный этап, в ходе которого глюкоза фосфорилируется и расщепляется на две молекулы фосфотриоз. Эта серия реакций протекает с использованием 2 молекул АТФ.
Этап, сопряжённый с синтезом АТФ. В результате этой серии реакций фосфотриозы превращаются в пируват. Энергия, высвобождающаяся на этом этапе, используется для синтеза 10 моль АТФ.
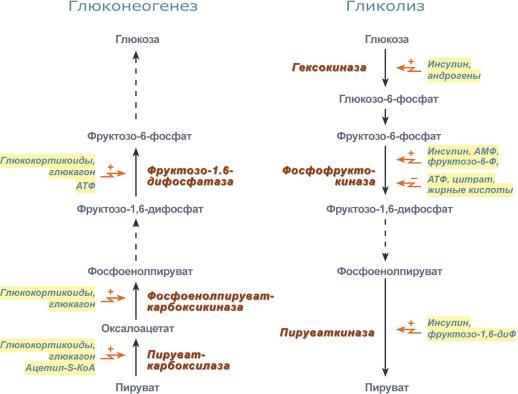
10…… Глюконеогенез — процесс образования в печени и отчасти в корковом веществе почек (около 10 %) молекул глюкозы из молекул других органических соединений — источников энергии, например свободных аминокислот, молочной кислоты, глицерина. Свободные жирные кислоты у млекопитающих для глюконеогенеза не используются.
Стадии глюконеогенеза
1Превращение пирувата в оксалоацетат (фермент пируваткарбоксилаза)
2Превращение оксалоацетата в фосфоенолпируват (фермент фосфоенолпируваткарбоксикиназа)
3Превращение фруктозо-1,6-дифосфата в фруктозо-6-фосфат (фермент фруктозо-1,6-дифосфатаза)
4Превращение глюкозо-6-фосфата в глюкозу (фермент глюкозо-6-фосфатаза)
Суммарное уравнение глюконеогенеза2 Пируват + 4 АТФ + 2 ГТФ + 2 (NADH + Н+)+ 4 Н20 → Глюкоза + 4 АДФ + 2 ГДФ + 6 H3PO4 + 2 NAD+
Роль в организме
При голодании в организме человека активно используются запасы питательных веществ (гликоген, жирные кислоты). Они расщепляются до аминокислот, кетокислот и других неуглеводных соединений. Большая часть этих соединений не выводится из организма, а подвергаются реутилизации. Вещества транспортируются кровью в печень из других тканей, и используются в глюконеогенезе для синтеза глюкозы — основного источника энергии в организме. Таким образом при истощении запасов организма, глюконеогенез является основным поставщиком энергетических субстратов.
11…..Регуляция гликолиза
В печени кроме пары противоположных процессов синтеза и распада гликогена, свойственной многим клеткам, есть и другая пара противоположных процессов — гликолиз и глюконеогенез (в других органах — только гликолиз). Как и в случаесинтеза и распада гликогена, регуляция гликолиза и глюконеогенеза связана с ритмом питания, с гормонами инсулином и глюкагоном и с фосфорилированием-де-фосфорилированием ферментов. Переключение печени с гликолиза на глюконеогенез и обратно происходит главным образом в результате изменений активности ферментов, образующих I и II субстратные циклы. Глюконеогенез включается в постабсорбтивном состоянии, когда концентрация глюкагона в крови повышена и, следовательно, активирована аденилатциклазная система и протеинкиназа А.
В регуляции I цикла основная роль принадлежит пируваткиназе, фосфорилиро-ванная форма которой неактивна, а дефосфорилированная — активна (рис. 9.31). Следовательно, гликолитическая реакция ФЕП -* пируват ускоряется при пищеварении и замедляется в постабсорбтивном состоянии. Что касается реакций этого цикла, связанных с глюконеогенезом (пируват -+ оксалоацетат -+ ФЕП), то, по всей вероятности, они с определенной скоростью происходят при любых состояниях. Это может быть связано с необходимостью поддержания в клетке определенной концентрации оксалоацетата, поскольку он участвует во многих важных процессах, включая цитратный цикл.
. 
13…..
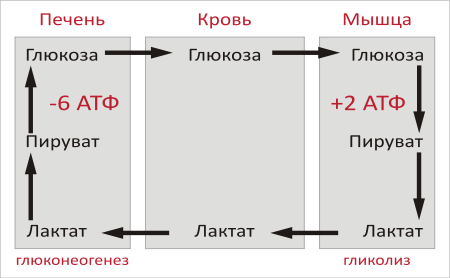
Цикл Кори — совокупность биохимических ферментативных процессов транспорта лактата из мышц в печень, и дальнейшего синтеза глюкозы из лактата, катализируемое ферментами глюконеогенеза.
При интенсивной мышечной работе, а также в условиях отсутствия или недостаточного числа митохондрий (например, в эритроцитах или мышцах) глюкоза вступает на путь анаэробного гликолиза с образованием лактата.Лактат не может далее окисляться, он накапливается (при его накоплении в мышцах раздражаются чувствительные нервные окончания, что вызывает характерное жжение в мышцах). С током крови лактат поступает в печень. Печень является основным местом скопления ферментов глюконеогенеза (синтез глюкозы из неуглеводныхсоеднений), и лактат идет на синтез глюкозы.
Реакция превращения лактата в пируват катализируется лактатдегидрогеназой, далее пируват подвергается окислительному декарбоксилированию или может подвергаться брожению.
14….. ПЕНТОЗОФОСФАТНЫЙ ПУТЬ ПРЕВРАЩЕНИЯ ГЛЮКОЗЫ
Пентозофосфатный путь, называемый также гексомонофосфатным шунтом, служит альтернативным путём окисления глюкозо-6-фосфата. Пентозофосфатный путь состоит из 2 фаз (частей) - окислительной и неокислительной.
В окислительной фазе глюкозо-6-фосфат необратимо окисляется в пентозу - рибулозо-5-фосфат, и образуется восстановленный NADPH.
В неокислительной фазе рибулозо-5-фосфат обратимо превращается в рибозо-5-фосфат и метаболиты гликолиза.
Пентозофосфатный путь обеспечивает клетки рибозой для синтеза пуриновых и пиримидиновых нуклеотидов и гидрированнымко-ферментом NADPH, который используется в восстановительных процессах.
Суммарное уравнение пентозофосфатного пути выражается следующим образом:
3 Глюкозо-6-фосфат + 6 NADP+ → 3 СО2 + 6 (NADPH + Н+) + 2 Фруктозо-6-фосфат + Глицеральдегид- 3 -фосфат.
Ферменты пентозофосфатного пути, так же, как и ферменты гликолиза, локализованы в цитозоле.
Наиболее активно Пентозофосфатный путь протекает в жировой ткани, печени, коре надпочечников, эритроцитах, молочной железе в период лактации, семенниках.
15…. Нарушения углеводного обмена при недостаточности инсулина.
Углеводный обмен при сахарном диабете характеризуется следующими особенностями:
1. резко снижен синтез глюкокиназы, которая при диабете почти полностью исчезает из печени, что ведет к уменьшению образования глюкозо-6-фосфата в клетках печени. Этот момент наряду со сниженным синтезом гликогенсинтетазы обусловливает резкое замедление синтеза гликогена. Происходит обеднение печени гликогеном. При недостатке глюкозо-6-фосфата тормозится пентозофосфатный цикл;
2. активность глюкозо-6-фосфатазы резко возрастает, поэтому глюкозо-6-фосфат дефосфорилируется и поступает в кровь в виде глюкозы;
3. тормозится переход глюкозы в жир;
4. понижается прохождение глюкозы через клеточные мембраны, она плохо усваивается тканями;
5. резко ускоряется глюконеогенез - образование глюкозы из лактата, пирувата, аминокислот жирных кислот и других продуктов неуглеводного обмена. Ускорение глюконеогенеза при сахарном диабете обусловлено отсутствием подавляющего влияния (супрессии) инсулина на ферменты, обеспечивающие глюконеогенез в клетках печени и почек: пируваткарбоксилазу, глюкозо-6-фосфатазу и др.
Тким образом, при сахарном диабете имеют место избыточная продукция и недостаточное использование глюкозы тканями, вследствие чего возникает гипергликемия. Содержание сахара в крови при тяжелых формах может достигать 4-5 г/л (400-500 мг%) и выше. При этом резко возрастает осмотическое давление крови, что ведет к обезвоживанию клеток организма. В связи с обезвоживанием глубоко нарушаются функции центральной нервной системы (гиперосмолярная кома).
Сахарная кривая при диабете по сравнению с таковой у здоровых значительно растянута во времени. Значение гипергликемии в патогенезе заболевания двояко. Она играет адаптивную роль, так как при ней тормозится распад гликогена и частично усиливается его синтез. При гипергликемии глюкоза лучше проникает в ткани и они не испытывают резкого недостатка углеводов. Гипергликемия имеет и отрицательное значение. При ней повышается концентрация глюко- и мукопротеидов, которые легко выпадают в соединительной ткани, способствуя образованию гиалина. Поэтому для сахарного диабета характерно раннее поражение сосудов атеросклерозом. Атеросклеротический процесс захватывает коронарные сосуды сердца (коронарная недостаточность), сосуды почек (гломерулонефриты) и др.
В пожилом возрасте сахарный диабет может сочетаться с гипертонической болезнью.
При повышении содержания сахара в крови до 1,6-2,0 г/л (160-200 мг%) и выше он начинает переходить в окончательную мочу - возникает глюкозурия.
Глюкозурия. В норме глюкоза содержится в провизорной моче. В канальцах она реабсорбируется в виде глюкозофосфата, для образования которого необходима гексокиназа, и после дефосфорилирования поступает в кровь. Таким образом, в окончательной моче сахара в нормальных условиях не содержится. При диабете процессы фосфорилирования и дефосфорилирования глюкозы в канальцах почек не справляются с избытком глюкозы в первичной моче. Развивается глюкозурия. При тяжелых формах сахарного диабета содержание сахара в моче может достигать 8-10%. Осмотическое давление мочи повышено; в связи с этим в окончательную мочу переходит много воды. Суточный диурез возрастает до 5-10 л и более (полиурия). Развивается обезвоживание организма и как следствие его - усиленная жажда (полидипсия).
ЭНЕРГЕТИЧЕСКИЙ ОБМЕН!!!!!!!!!!!!!!!!!!!!!!!!
1……. Обмен веществ или метаболизм представляет собой высокоинтегрированый и целенаправленный процесс, в котором участвует много ферментативных систем и который обеспечен сложнейшей регуляцией на разных уровнях.
С точки зрения термодинамики живые организмы представляют собой открытые системы, поскольку они обмениваются с окружающей средой, как энергией, так и веществом, и при этом преобразуют и то и другое. При наблюдении в течение определённого отрезка времени в химическом составе организма определённых изменений не происходит. Но это не значит что химические вещества, составляющие организм не подвергаются ни каким изменениям. Напротив они постоянно и достаточно интенсивно обновляются. Это потому что скорость переноса веществ и энергии из среды в организм точно уравновешивается скоростью переноса из организма в среду.
превращений веществ, которые происходят в организме начиная с момента поступления переваренных пищевых веществ в кровь и до момента выделения конечных продуктов обмена из организма – называют промежуточным метаболизмом (обменом веществ). Промежуточный метаболизм может быть разделён на два процесса: катаболизм (диссимиляция) и анаболизм (ассимиляция). Катаболизм – это ферментативное расщепление сравнительно крупных органических молекул осуществляемое у высших организмов, как правило, окислительным путём. Катаболизм сопровождается освобождением энергии, заключенной в сложных структурах крупных органических молекул и запасанием её в форме фосфатных связей АТФ. Анаболизм – это ферментативный синтез из более простых соединений крупномолекулярных клеточных компонентов, таких как полисахариды, нуклеиновые кислоты, белки, липиды, а также некоторых их предшественников. Анаболические процессы протекают с потреблением энергии. Катаболизм и анаболизм происходят в клетках одновременно и неразрывно связаны друг с другом. По существу их следует рассматривать не как два отдельных процесса, а как две стороны одного общего процесса – метаболизма, в котором превращение веществ теснейшим образом переплетены с превращением энергии.
Катаболические и анаболические реакции происходят одновременно, но в разных частях клетки. Например, окисление жирных кислот осуществляется с помощью набора ферментов локализованных в митохондриях, тогда как синтез жирных кислот катализирует другая система ферментов, локализующая в цитозоле. Именно благодаря разной локализации катаболические и анаболические процессы в клетке могут протекать одновременно.
22222….. Клеточное или тканевое дыхание — совокупность биохимических реакций, протекающих в клетках живых организмов, в ходе которых происходит окисление углеводов, липидов и аминокислот до углекислого газа и воды. Высвобожденная энергия запасается в химических связях макроэргических соединений (АТФ и др.) и может быть использована по мере необходимости. Входит в группу процессов катаболизма. О физиологических процессах транспортировки к клеткам многоклеточных организмов кислорода и удалению от них углекислого газа
Гликолиз — путь ферментативного расщепления глюкозы — является общим практически для всех живых организмов процессом. У аэробов он предшествует собственно клеточному дыханию, у анаэробов завершается брожением. Сам по себе гликолиз является полностью анаэробным процессом и для осуществления не требует присутствия кислорода.
Первый его этап протекает с расходом энергии 2 молекул АТФ и включает в себя расщепление молекулы глюкозы на 2 молекулы глицеральдегид-3-фосфата. На втором этапе происходит НАД-зависимое окисление глицеральдегид-3-фосфата, сопровождающееся субстратным фосфорилированием, то есть присоединением к молекуле остатка фосфорной кислоты и формированием в ней макроэргической связи, после которого остаток переносится на АДФ с образованием АТФ.
Таким образом, уравнение гликолиза имеет следующий вид:
Глюкоза + 2НАД+ + 4АДФ + 2АТФ + 2Фн = 2ПВК + 2НАД∙Н + 2 АДФ + 4АТФ + 2H2O + 4Н+.
Сократив АТФ и АДФ из левой и правой частей уравнения реакции, получим:
Глюкоза + 2НАД+ + 2АДФ + 2Фн = 2НАД∙Н + 2ПВК + 2АТФ + 2H2O + 2Н+.
Ацетил-КоА под действием цитратсинтазы передаёт ацетильную группу оксалоацетату с образованием лимонной кислоты, которая поcтупает в цикл трикарбоновых кислот (цикл Кребса). В ходе одного оборота цикла лимонная кислота несколько раз дегидрируется и дважды декарбоксилируется с регенерацией оксалоацетата и образованием одной молекулы ГТФ (способом субстратного фосфорилирования), трёх НАДН и ФАДН2.
Суммарное уравнение реакций:
Ацетил-КоА + 3НАД+ + ФАД + ГДФ + Фн + 2H2O + КоА-SH = 2КоА-SH + 3НАДH + 3H+ + ФАДН2 + ГТФ + 2CO2
У эукариот ферменты цикла находятся в свободном состоянии в матриксе митохондрий, только сукцинатдегидрогеназавстроена во внутреннюю митохондриальную мембрану.
33333… Окислительноефосфорилирование — один из важнейших компонентов клеточного дыхания, приводящего к получению энергии в виде АТФ. Субстратами окислительного фосфорилирования служат продукты расщепления органических соединений — белки, жиры и углеводы.
Предварительно сложные углеводы расщепляются до простых, вплоть до образования глюкозы. Глюкоза является универсальным субстратом в процессе клеточного дыхания. Окисление глюкозы подразделяется на 3 этапа:
1гликолиз;
2окислительное декарбоксилирование или цикл Кребса;
3 окислительное фосфорилирование.
Работа АТФ-синтазы
Процесс окислительногофосфорилирования осуществляется пятым комплексом дыхательной цепи митохондрий — Протонная АТФ-синтаза, состоящая из 9 субъединиц 5 типов:
-3 субъединицы (γ,δ,ε) способствуют целостности АТФ-синтазы
-β субъединица является основной функциональной единицей. Она имеет 3 конформации:
-L-конформация — присоединяет АДФ и Фосфат (поступают в митохондрию из цитоплазмы с помощью специальных переносчиков)
-Т-конформация — к АДФ присоединяется фосфат и образуется АТФ
-О-конформация — АТФ отщепляется от β-субъединицы и переходит на α-субъединицу.
Для того, чтобы субъединица изменила конформацию необходим протон водорода, так как конформация меняется 3 раза необходимо 3 протона водорода. Протоны перекачиваются из межмембранного пространства митохондрии под действием электрохимического потенциала.
44444…. Дыхательная цепь — это главная система превращения энергии в митохондриях. В ней последовательно окисляются и восстановливаются элементы дыхательной цепи, в результате чего высвобождается небольшими порциями энергия. За счет этой энергии в трех точках этой цепи из АДФ и фосфата образуется АТФ. Поэтому говорят, что окисление (перенос электронов) сопряжено с фосфорилированием (АДФ + Фн → АТФ), т.е. происходит процесс окислительного фосфорилирования.
В результате многократной оборачиваемости субстратов в цикле Кребса полностью окисляются поступившие продукты первичного гликолитического окисления. Затем в цепи окислительного фосфорилирования энергия, освободившаяся при окислении, максимально используется для синтеза АТФ.
Было высказано предположение, что выделяющаяся при транспорте электронов энергия запасается в виде градиента протонов на мембране. При этом на внешней поверхности внутренней мембраны митохондрий возникает повышенная концентрация положительно заряженных ионов водорода. Возникший при этом протонный градиент является движущей силой в синтезе АТФ.
55555 ….
Перенос электронов от NADH к О2 включает ряд переносчиков, которые локализованы во внутренней мембране митохондрий. За исключением убихинона и цитохрома С, это сложные белковые комплексы.
NADH-дегидрогеназа (NADH-Q-редуктаза, комплекс I) состоит из нескольких полипептидных цепей. Роль простетической группы играет FMN. Единственный субстрат фермента - NADH, с которого 2 электрона и протон переносятся на FMN с образованием FMNH2. Второй протон поглощается из матрикса. Реакция протекает по уравнению:
NADH + Н+ + Е (FMN) → NAD+ + Е (FMNH2)
С FMNH2 электроны переносятся затем на ряд железо-серных белков (FeS), играющих роль второй простетической группы в молекуле NADH-дегидрогеназы. Атомы железа в этих белках (негемовое железо) собраны в несколько групп, так называемых железо-серных центров. FeS-центры входят в состав многих белков (флавопротеинов, цитохромов), участвующих в окислительно-восстановительных реакциях. Известны 3 типа FeS-центров (FeS, Fe2S2, Fe4S4), в которых атом железа связан с атомом серы остатков цистеина или неорганической серы. Строение железо-серных центров показано на рис. 6-7.
NADH-дегидрогеназа содержит несколько центров типа Fe2S2 и Fe4S4 Атомы железа в таких центрах могут принимать и отдавать электроны поочерёдно, переходя в ферро- (Fe2+) и ферри- (Fe3+) состояния. От железо-серных центров электроны переносятся на кофермент Q (убихинон)
66666… Обозначение этого жирорастворимого хинона происходит от первой буквы английского названия хинона (quinone), а название убихинон отражает его широкую распространённость в природе (ubiquitous - вездесущий). Молекулы убихинона в зависимости от источника, из которого они выделены, различаются длиной углеводородной цепи, которая у млекопитающих содержит 10 изопреноидных звеньев и обозначается как Q10. В процессе переноса электронов с NADH-дегидрогеназы через FeS на убихинон он обратимо превращается в гидрохинон. Убихинон выполняет коллекторную функцию, присоединяя электроны от NADH-дегидрогеназы и других флавинзависимыхдегидрогеназ, в частности, от сукцинат-дегидрогеназы. Убихинон участвует в реакциях типа:
Е (FMNH2) + Q → Е (FMN) + QH2.
В зависимости от способности поглощать свет в определённой части спектра все цитохромы делят на группы а, b, с. Внутри каждой группы отдельные виды с уникальными спектральными свойствами обозначают цифровыми индексами (b, b1, b2 и т.д.).
Структурные особенности разных видов цитохромов определяют различие в их окислительно-восстановительных потенциалах. В ЦПЭ участвуют 5 типов цитохромов (а, а3, b, с, с1). За исключением цитохрома с, все цитохромы находятся во внутренней мембране митохондрий в виде сложных белковых комплексов (табл. 6-4).
QН2-дегидрогеназа (коэнзим Q-цитохром с-ре-уктаза, комплекс III) состоит из 2 типов цитохромов (b1 и b2) и цитохрома с1. QН2-дегидрогеназа переносит электроны от убихинола на цитохром с. Внутри комплекса III электроны передаются от цитохромов b на FeS-центры, на цитохром с1, а затем на цитохром с. Группы тема, подобно FeS-центрам, переносят только по одному электрону. Таким образом, от молекулы QH2 2 электрона переносятся на 2 молекулы цитохрома b. В качестве промежуточного продукта в этих реакциях переноса электронов возможно образование свободного радикала се-михинона. В цитохромах типа b гем не связан ковалентно с белком, а в цитохромах с1 и с он присоединяется к белку при помощи тиоэфирных связей (рис. 6-9). Эти связи образуются путём присоединения 2 цистеиновых остатков к винильным группам гема.
Цитохром с - периферический водорастворимый мембранный белок с молекулярной массой 12 500 Д, имеющий одну полипептидную цепь из 100 аминокислотных остатков, и молекулу гема, ковалентно связанную с полипептидом.
Цитохромоксидаза (комплекс IV) состоит из 2 цитохромов типа аа3 каждый из которых имеет центр связывания с кислородом. Цитохромы а и а3 имеют характерную железопорфириновую простетическую группу, называемую гемом А и отличающуюся от гемацитохромов с и c1 (рис. 6-10). Он содержит формильную группу вместо одной из метальных групп и углеводородную цепь вместо одной из винильных групп.
Другая особенность комплекса а-а3 - наличие в нём ионов меди, связанных с белковой частью в так называемых CuA-центрах. Перенос электронов комплексом а-а3 включает реакции:
Cu+ ↔ Cu2+ + e,
Fe2+ ↔ Fe3+ + e
777 … Строение АТФ-синтазы и синтез АТФ
АТФ-синтаза (Н+-АТФ-аза) - интегральный белок внутренней мембраны митохондрий. Он расположен в непосредственной близости к дыхательной цепи. АТФ-синтаза состоит из 2 белковых комплексов, обозначаемых как F0 и F1 (рис. 6-15).
Гидрофобный комплекс F0 погружён в мембрану. Он служит основанием, которое фиксирует АТФ-синтазу в мембране. Комплекс F0 состоит из нескольких субъединиц, образующих канал, по которому протоны переносятся в матрикс.
А - F0 и F1 - комплексы АТФ-синтазы, В состав F0 входят полипептидные цепи, которые образуют канал, пронизывающий мембрану насквозь. По этому каналу протоны возвращаются в матрикс из межмембранного пространства; белок F1 выступает в матрикс с внутренней стороны мембраны и содержит 9 субъединиц, 6 из которых образуют 3 пары α и β ("головка"), прикрывающие стержневую часть, которая состоит из 3 субъединиц γ, δ и ε. γ и ε подвижны и образуют стержень, вращающийся внутри неподвижной головки и связанный с комплексом F0. В активных центрах, образованных парами субъединиц α и β, происходит связывание АДФ, неорганического фосфата (Рi) и АТФ. Б - Каталитический цикл синтеза АТФ включает 3 фазы, каждая из которых проходит поочерёдно в 3 активных центрах: 1 - связывание АДФ и Н3РО4; 2 - образование фосфоангидридной св
|
| Поделиться: |
Поиск по сайту
Все права принадлежать их авторам. Данный сайт не претендует на авторства, а предоставляет бесплатное использование.
Дата создания страницы: 2022-10-12 Нарушение авторских прав и Нарушение персональных данных