В гепатоцитах содержатся практически все ферменты, участвующие в метаболизме липидов. Поэтому паренхиматозные клетки печени в значительной степени контролируют соотношение между потреблением и синтезом липидов в организме. Катаболизм липидов в клетках печени протекает главным образом в митохондриях и лизосомах, биосинтез - в цитозоле и эндоплазматическом ретикулуме. Ключевым метаболитом липидного обмена в печени является ацетил-КоА, главные пути образования и использования которого показаны на рисунке 5.

Рисунок 5. Образование и использование ацетил-КоА в печени.
31.3.1. Метаболизм жирных кислот в печени. Пищевые жиры в виде хиломикронов поступают в печень через систему печёночной артерии. Под действием липопротеинлипазы, находящейся в эндотелии капилляров, они расщепляются до жирных кислот и глицерола. Жирные кислоты, проникающие в гепатоциты, могут подвергаться окислению, модификации (укорочению или удлинению углеродной цепи, образованию двойных связей) и использоваться для синтеза эндогенных триацилглицеролов и фосфолипидов.
31.3.2. Синтез кетоновых тел. При β-окислении жирных кислот в митохондриях печени образуется ацетил-КоА, подвергающийся дальнейшему окислению в цикле Кребса. Если в клетках печени имеется дефицит оксалоацетата (например, при голодании, сахарном диабете), то происходит конденсация ацетильных групп с образованием кетоновых тел (ацетоацетат,β-гидроксибутират, ацетон). Эти вещества могут служить энергетическими субстратами в других тканях организма (скелетные мышцы, миокард, почки, при длительном голодании - головной мозг). Печень не утилизирует кетоновые тела. При избытке кетоновых тел в крови развивается метаболический ацидоз. Схема образования кетоновых тел - на рисунке 6.

Рисунок 6. Синтез кетоновых тел в митохондриях печени.
31.3.3. Образование и пути использования фосфатидной кислоты. Общим предшественником триацилглицеролов и фосфолипидов в печени является фосфатидная кислота. Она синтезируется из глицерол-3-фосфата и двух ацил-КоА - активных форм жирных кислот (рисунок 7). Глицерол-3-фосфат может образоваться либо из диоксиацетонфосфата (метаболит гликолиза), либо из свободного глицерола (продукт липолиза).

Рисунок 7. Образование фосфатидной кислоты (схема).
Для синтеза фосфолипидов (фосфатидилхолина) из фосфатидной кислоты необходимо поступление с пищей достаточного количества липотропных факторов (веществ, препятствующих развитию жировой дистрофии печени). К этим факторам относятся холин, метионин, витамин В12, фолиевая кислота и некоторые другие вещества. Фосфолипиды включаются в состав липопротеиновых комплексов и принимают участие в транспорте липидов, синтезированных в гепатоцитах, в другие ткани и органы. Недостаток липотропных факторов (при злоупотреблении жирной пищей, хроническом алкоголизме, сахарном диабете) способствует тому, что фосфатидная кислота используется для синтеза триацилглицеролов (нерастворимых в воде). Нарушение образования липопротеинов приводит к тому, что избыток ТАГ накапливается в клетках печени (жировая дистрофия) и функция этого органа нарушается. Пути использования фосфатидной кислоты в гепатоцитах и роль липотропных факторов показаны на рисунке 8.
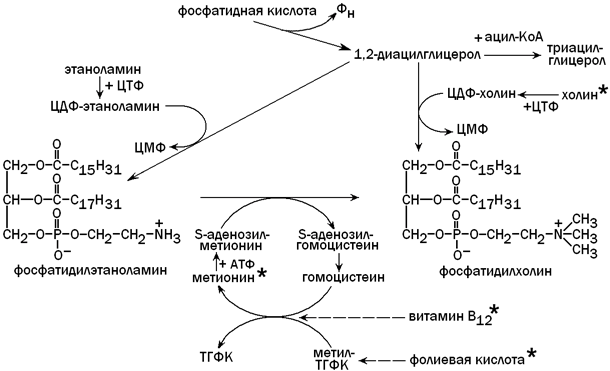
Рисунок 8. Использование фосфатидной кислоты для синтеза триацилглицеролов и фосфолипидов. Липотропные факторы обозначены знаком *.
31.3.4. Образование холестерола. Печень является основным местом синтеза эндогенного холестерола. Это соединение необходимо для построения клеточных мембран, является предшественником желчных кислот, стероидных гормонов, витамина Д3. Первые две реакции синтеза холестерола напоминают синтез кетоновых тел, но протекают в цитоплазме гепатоцита. Ключевой фермент синтеза холестерола - β-гидрокси-β-метилглутарил-КоА-редуктаза (ГМГ-КоА-редуктаза) ингибируется избытком холестерола и желчными кислотами по принципу отрицательной обратной связи (рисунок 9).

Рисунок 9. Синтез холестерола в печени и его регуляция.
31.3.5. Образование липопротеинов. Липопротеины - белково-липидные комплексы, в состав которых входят фосфолипиды, триацилглицеролы, холестерол и его эфиры, а также белки (апопротеины). Липопротеины транспортируют нерастворимые в воде липиды к тканям. В гепатоцитах образуются два класса липопротеинов - липопротеины высокой плотности (ЛПВП) и липопротеины очень низкой плотности (ЛПОНП).
93. Роль печени в азотистом обмене. Пути использования фонда аминокислот в печени. Особенности в детском возрасте.
Печень является органом, регулирующим поступление азотистых веществ в организм и их выведение. В периферических тканях постоянно протекают реакции биосинтеза с использованием свободных аминокислот, либо выделение их в кровь при распаде тканевых белков. Несмотря на это, уровень белков и свободных аминокислот в плазме крови остаётся постоянным. Это происходит благодаря тому, что в клетках печени имеется уникальный набор ферментов, катализирующих специфические реакции обмена белков.
31.4.1. Пути использования аминокислот в печени. После приёма белковой пищи в клетки печени по воротной вене поступает большое количество аминокислот. Эти соединения могут претерпевать в печени ряд превращений, прежде чем поступить в общий кровоток. К этим реакциям относятся (рисунок 10):
а) использование аминокислот для синтеза белков;
б) трансаминирование - путь синтеза заменимых аминокислот; осуществляет также взаимосвязь обмена аминокислот с глюконеогенезом и общим путём катаболизма;
в) дезаминирование - образование α-кетокислот и аммиака;
г) синтез мочевины - путь обезвреживания аммиака (схему см. в разделе "Обмен белков");
д) синтез небелковых азотсодержащих веществ (холина, креатина, никотинамида, нуклеотидов и т.д.).

Рисунок 10. Обмен аминокислот в печени (схема).
31.4.2. Биосинтез белков. В клетках печени синтезируются многие белки плазмы крови: альбумины (около 12 г в сутки), большинство α- и β-глобулинов, в том числе транспортные белки (ферритин, церулоплазмин, транскортин, ретинолсвязывающий белок и др.). Многие факторы свёртывания крови (фибриноген, протромбин, проконвертин, проакцелерин и др.) также синтезируются в печени.
Компартментализация метаболических процессов в печени. Регуляция направления потока метаболитов через мембраны внутриклеточных (субклеточных) структур. Значение в интеграции обмена веществ.
Клетка - сложнофункциональная система, регулирующая своё жизнеобеспечение. Многообразие функций клетки обеспечивается пространственной и временной (в первую очередь, в зависимости от ритма питания) регуляцией определённых метаболических путей. Пространственная регуляция связана со строгой локализацией определённых ферментов в различных
Таблица 2-3. Типы метаболических путей
| Схема | Название | Пример |
| А→В→С→D→Е | Линейный | Гликолиз |

| Разветвлённый | Синтез нуклеотидов |

| Циклический | Цикл трикарбоновых кислот |
| Синтез мочевины | ||

| Спиральный | β-окисление жирных кислот |
органеллах. Так, в ядре находятся ферменты, связанные с синтезом молекул ДНК и РНК, в цитоплазме - ферменты гликолиза, в лизосомах - гидролитические ферменты, в матриксе митохондрий - ферменты ЦТК, во внутренней мембране митохондрий - ферменты цепи переноса электронов и т.д. (рис. 2-29). Такая субклеточная локализация ферментов способствует упорядоченности биохимических процессов и увеличивает скорость обмена веществ.
Роль печени в обезвреживании ксенобиотиков. Механизмы обезвреживания веществ в печени. Стадии (фазы) химической модификации. Роль реакций конъюгации в детоксикации продуктов обмена и лекарственных препаратов (примеры). Метаболизм лекарств у детей раннего возраста.
Основным представителем неспецифических транспортных систем крови является сывороточный альбумин. Этот белок может связывать практически все экзогенные и эндогенные низкомолекулярные вещества, что в значительной степени обусловлено его способностью легко изменять конформацию своей молекулы и большим количеством гидрофобных участков в молекуле.
Различные вещества связываются с альбумином крови нековалентными связями: водородными, ионными, гидрофобными. При этом различные группы веществ взаимодействуют с определёнными группировками альбумина, вызывая характерные изменения конформации его молекулы. Существует представление, что вещества, которые прочно связываются с белками крови, обычно экскретируются печенью с желчью, а вещества, образующие слабые комплексы с белками, выводятся почками с мочой.
Связывание лекарств с белками крови снижает скорость их утилизации в тканях и создаёт определённый резерв их в кровяном русле. Интересно отметить, что у больных с гипоальбуминемией чаще встречаются побочные реакции при введении лекарств вследствие нарушения транспорта их к клеткам-мишеням.
33.4.3. Внутриклеточные транспортные системы. В цитоплазме клеток печени и других органов имеются белки-носители, которые ранее обозначались как Y- и Z-белки или лигандины. В настоящее время установлено, что эти белки являются различными изоферментами глутатион-S-трансферазы. Эти белки связывают большое количество различных соединений: билирубин, жирные кислоты, тироксин, стероиды, канцерогены, антибиотики (бензилпенициллин, цефазолин, хлорамфеникол, гентамицин). Известно, что эти трансферазы играют роль в транспорте указанных веществ из плазмы крови через гепатоциты в печень.