Организм осуществляет тонкое регулирование напряжения О2 и СО2 в крови – их содержание остается относительно постоянным, несмотря на колебания количества доступного кислорода и потребности в нем, которая во время интенсивной мышечной работы может увеличиваться в 20 раз. Частота и глубина дыхания регулируются дыхательным центром, нейроны которого расположены в различных отделах ЦНС; главными из них являются продолговатый мозг и мост. Дыхательный центр по соответствующим нервам ритмично посылает к диафрагме и межреберным мышцам импульсы, которые вызывают дыхательные движения. В основе своей ритм дыхания является непроизвольным, но может изменяться в некоторых пределах высшими центрами головного мозга, что свидетельствует о возможности произвольного влияния на нижележащие отделы дыхательного центра.
Саморегуляция вдоха и выдоха
Локализация дыхательного центра. В продолговатом мозге находится главная часть дыхательного центра. Об этом свидетельствуют исследования П.Флуранса (1794-1867) [6;217]. Он обнаружил, что разрушение медиальной части продолговатого мозга в нижнем углу ромбовидной ямки ведет к полной остановке дыхания. Позже (1885) Н. А. Миславский [7;39] установил наличие двух структур, ответственных за вдох и выдох.
Мост играет важную роль в регуляции продолжительности фаз вдоха, выдоха и паузы между ними.
Мотонейроны спинного мозга получают импульсы от нейронов продолговатого мозга и посылают их к дыхательным мышцам по диафрагмальному и межреберным нервам. Центр диафрагмальных нервов находится в основном в 3-4-м шейных сегментах спинного мозга. Центры межреберных нервов, иннервирующих мускулатуру грудной клетки, локализуются в грудном отделе спинного мозга (4 – 10 сегменты), иннервация мышц живота осуществляется Th4 – L3 – сегментами.
В регуляции дыхания принимают участие также средний мозг, гипоталамус, лимбико-ретикулярный комплекс, кора большого мозга.
Дыхательные нейроны (нейроны, которые возбуждаются в различные фазы дыхательного цикла) обнаружены почти на всем протяжении продолговатого мозга. Однако в обеих половинах продолговатого мозга есть участки ретикулярной формации, где имеются основные скопления дыхательных нейронов. В правой и левой половинах продолговатого мозга имеется по два таких скопления - дорсальное и вентральное. Они локализуются вблизи задвижки (obex), которая расположена у нижнего угла ромбовидной ямки.
Нейронная организация автоматии дыхательного центра. Под автоматией дыхательного центра понимают циркуляцию возбуждения в его нейронах, обеспечивающую саморегуляцию вдоха и выдоха. Взаимодействие нейронов дыхательного центра заключается в следующем. Ритмическая смена вдоха и выдоха (постоянное их чередование) обеспечивается циркуляцией возбуждения вдыхательных нейронах продолговатого мозга- главной части дыхательного центра, а также взаимодействием импульсации нейронов продолговатого мозга с импульсацией дыхательных нейронов моста и рефлексогенных зон, главной из которых является легочная (механорецепторы). При этом эфферентные импульсы ритмично поступают по диафрагмальному и межреберным нервам к дыхательным мышцам, что ведет к их сокращению (вдох). Прекращение импульсации сопровождается расслаблением дыхательной мускулатуры (выдох) [8;238].
Цикл дыхания у человека состоит из вдоха, выдоха и паузы. С учетом этого дыхательные нейроны классифицируют на группы, основными из которых являются: 1) ранние инспираторные и экспираторные нейроны; 2) поздние инспираторные и экспираторные нейроны; 3) полные инспираторные и экспираторные нейроны.
Большинство экспираторных нейронов являются антиинспираторными, и только часть из них посылают свои импульсы к мышцам выдоха. Они возбуждаются под влиянием афферентной импульсации блуждающих нервов и нейронов моста. Большинство инспираторных нейронов обладают непрерывной спонтанной импульсной активностью, которая преобразуется в фазную благодаря тормозным реципрокным влияниям экспираторных и поздних инспираторных нейронов [9;218].
Каждый дыхательный цикл начинается с возбуждения ранних инспираторных нейронов. Затем возбуждение переходит на полные инспираторные нейроны. В процессе циркуляции возбуждения импульсы по возвратным связям поступают к предшествующим нейронам и тормозят их. Полные инспираторные и экспираторные нейроны по нисходящим путям посылают импульсы к мотонейронам спинного мозга, иннервирующим дыхательную мускулатуру (рис. 6.1).
Представленная схема саморегуляции вдоха и выдоха на рис. 6.1 не отражает ряд известных в настоящее время процессов взаимодействия дыхательных нейронов моста, продолговатого мозга и афферентных импульсов от рефлексогенных зон, но она хорошо иллюстрирует принципиальную структуру механизмов саморегуляции вдоха и выдоха. Показаны три источника импульсов, обеспечивающих смену вдоха на выдох, торможение инспираторных нейронов (Ир и И): от нейронов моста (М), от самих инспираторных нейронов (Ир и И) и от рецепторов легких, импульсация которых поступает по блуждающим нервам [10;201].
Роль рефлексогенных зон в регуляции вдоха и выдоха
Роль блуждающих нервов в регуляции вдоха и выдоха доказали Геринг и Брейер в опыте с раздуванием легких воздухом в различные фазы дыхательного цикла. Оказалось, что раздувание легких воздухом тормозит вдох, после чего наступает выдох. Уменьшение объема легких (забор воздуха) тормозит выдох, ускоряет вдох. После перерезки блуждающих нервов раздувание легких не изменяет характер дыхания – тормозной эффект отсутствует.
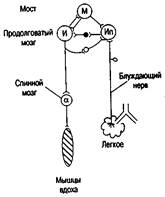
Рис. 2.1. Схема, отражающая основные процессы саморегуляции вдоха и выдоха. Нейроны: М — ретикулярной формации моста, И – инспираторные (их совокупность, кроме Ип), Ип - инспираторные поздние, а — мотонейроны спинного мозга, нейроны: черный - тормозящий, светлый - возбуждающий
Значение проприорецепторов дыхательных мышц в регуляции дыхания является таким же, как и для всей скелетной мускулатуры. Причем главную роль играют проприорецепторы (мышечные веретена и сухожильные рецепторы) межреберных мышц и мышц стенки живота, которые содержат большое количество этих рецепторов. Импульсация от проприорецепторов усиливает сокращение дыхательной мускулатуры и способствует смене вдоха на выдох.
Рецепторы верхних дыхательных путей — в основном солодовые, при своем возбуждении оказывают слабое тормозящее влияние на дыхание [11;320].
Раздражение обонятельных, рецепторов пахучими веществами в умеренной концентрации вызывает короткие вдохи – принюхивание. Сильное раздражение оболочек воздухоносных путей (пыль, едкие пары, например аммиака, и инородные тела) вызывает возбуждение окончаний тройничного нерва - возникает чихание, возможна остановка дыхания. Раздражение рецепторов гортани и трахеи сопровождается кашлем. Чихание, кашель, смыкание голосовых связок и сужение бронхов, препятствующие попаданию инородных частиц в нижние дыхательные пути,- это защитные рефлексы. К этой же категории относится и так называемый рефлекс ныряльщиков — рефлекторное апноэ при действии воды на область нижних носовых ходов.
Сильное возбуждение тепловых или Холодовых рецепторов кожи может привести к возбуждению дыхательного центра и усилению дыхания. Однако погружение человека в холодную воду тормозит выдох, и возникает затяжной вдох.
Повышение температуры тела при заболеваниях также сопровождается увеличением вентиляции легких [12;180].
Незначительное понижение температуры тела стимулирует дыхание, а глубокая гипотермия угнетает дыхательный центр.
Нейрогуморальная регуляция интенсивности дыхания
В опытах Фредерика и Холдена установлено, что главным стимулятором дыхания является СО2. Позже было показано, что увеличение содержания СО2 в альвеолах на 0,2% ведет к увеличению вентиляции легких на 100%. При снижении рН артериальной крови по сравнению с нормальным уровнем (7,4) вентиляция легких увеличивается. В случае возрастания рН выше нормы вентиляция уменьшается, хотя и в меньшей степени. Увеличение содержания СО2 в крови стимулирует дыхание как за счет снижения рН, так и непосредственным действием самого СО2.
Влияние СО2 и ионов Н+ на дыхание опосредованно, главным образом, их действием на особые структуры ствола мозга, обладающие хемочувствительностью (центральные хеморецепторы). Хеморецепторы, реагирующие на изменение газового состава крови, обнаружены в стенках сосудов только в двух областях – в дуге аорты и синокаротидной области (вне сосудов).
Снижение напряжения О2 в артериальной крови (гипоксемия) ниже 50-60 мм рт.ст. сопровождается увеличением вентиляции легких уже через 3-5 с. В норме такого сильного падения напряжения О2 не встречается, однако артериальная гипоксия может возникнуть при подъеме на высоту, при сердечно-легочной патологии. Сосудистые хеморецепторы (аортальные и синокаротидные) возбуждаются и при нормальном напряжении газов крови, их активность сильно возрастает при гипоксии и исчезает при дыхании чистым кислородом. Стимуляция дыхания при снижении напряжения О2 опосредована исключительно периферическими хеморецепторами. Аортальные и каротидные тельца возбуждаются (импульсация от них учащается) при повышении напряжения СО2 или при уменьшении рН. Однако влияние СО2 с хеморецепторов выражено меньше, нежели О2 [13;278].
Весьма важным является тот факт, что гипоксемия стимулирует дыхание значительно больше, если она сопровождается гиперкапнией, что наблюдается при физической работе: гипоксемия увеличивает реакцию на СО2. Однако при значительной гипоксемии вследствие снижения окислительного метаболизма уменьшается чувствительность центральных хеморецепторов. В этих условиях решающую роль в стимуляции дыхания играют сосудистые хеморецепторы, активность которых повышается, так как для них адекватным раздражителем является снижение напряжения кислорода в артериальной крови (аварийный механизм стимуляции дыхания). Таким образом, сосудистые хеморецепторы преимущественно реагируют на снижение в крови уровня кислорода, центральные хеморецепторы реагируют преимущественно на изменения в крови и спинномозговой жидкости рН и Рсо2.
Адаптация дыхания к движениям происходит путем различной интеграции объемов и емкостей общей емкости легких, легочной вентиляции, изменения степени равномерности вентиляции и диффузионной способности альвеолярной мембраны. Важное значение для эффективности дыхания при этом имеет координация функций дыхания и кровообращения. Обучение движениям приводит к специализированной интеграции дыхательной функции. Чем прочнее связь дыхания и движений, тем легче при прочих равных условиях выполняются движения. При этом сами дыхательные движения становятся как бы компонентами выученных движений.
ВЫВОДЫ
Итак, можно сделать такие выводы:
Физиологической основой общей выносливости для большинства современных видов профессиональной деятельности являются аэробные способности: они относительно малоспецифичны и мало зависят от вида выполняемых упражнений. Чем ниже мощность выполняемой работы и больше количество участвующих в ней мышц, тем в меньшей степени её результативность будет зависеть от совершенства двигательного навыка и больше – от аэробных возможностей, функциональные возможности вегетативных систем организма будут высокими при выполнении всех упражнений аэробной направленности. Именно поэтому выносливость к работе такой направленности имеет общий характер и её называют общей выносливостью.
Общая выносливость является основой высокой физической работоспособности, необходимой для успешной профессиональной деятельности. За счёт высокой мощности и устойчивости аэробных процессов быстрее восстанавливаются внутримышечные энергоресурсы и компенсируются неблагоприятные сдвиги во внутренней среде организма в процессе самой работы, обеспечивается переносимость высоких объёмов интенсивных силовых, скоростно-силовых физических нагрузок и координационно-сложных двигательных действий, ускоряется течение восстановительных процессов в периоды между тренировками.
При работе в несколько раз увеличивается объемная скорость кровотока, что обеспечивает доставку нужного количества кислорода к работающим мышцам и транспорт углекислого газа к альвеолярным капиллярам. При этом может возникать венозная гипоксемия (до 20—30% НbО2 вместо 60% НbО2 в покое). Если же усиливается неравномерность вентиляции и возникает недостаточная координация дыхания и кровотока в малом круге кровообращения, то возникает артериальная гипоксемия, достигающая при тяжелой и длительной работе 80% НbО2 и ниже. Эти явления отражают приспособительные механизмы повышения утилизации кислорода из крови для удовлетворения окислительных процессов в напряженно работающих мышцах.
Количество кислорода, необходимое для окислительных процессов, обеспечивающих ту или иную работу, называется кислородным запросом. Различают суммарный, или обший, кислородный запрос, т. е. количество кислорода, необходимое для выполнения всей работы, и минутный кислородный запрос, т. е. количество кислорода, потребляемое при данной работе в течение 1 мин.
Обычно развитие тренированности характеризуется процессом экономизации минутного и общего кислородного запроса от 7—8% до 30% и более к исходному уровню. Это происходит при увеличении эффективности выполняемой работы. При относительно равномерной работе (с колебаниями мощности в пределах ±3%), если она легко выполняется спортсменом, может наступить вскоре после начала работы равновесие между кислородным запросом и его удовлетворением, или истинное устойчивое состояние. При напряженной циклической работе, когда минутный кислородный запрос удовлетворяется не в полной мере (поскольку функции кровообращения и дыхания характеризуются предельно возможным уровнем транспорта кислорода), отмечается устойчивая величина потребления кислорода. Однако такая устойчивость потребления кислорода сопряжена с нарастанием кислородного долга, не проявляющегося по ходу работы. В этих случаях говорят о кажущемся устойчивом состоянии. Таким образом, истинное устойчивое состояние может поддерживаться длительное время, кажущееся же устойчивое состояние ограничено временем достижения предельно переносимой величины кислородного долга.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
1. Зимкина Н.В. Физиология человека. – М.: Просвещение, 1975. – 420 с.
2. Развитие выносливости спортом. – М.: Физкультура и спорт, 1989. –140 с.
3. Шабатура Н.Н., Матяш Н.Ю., мотузный В.А. Биология человека. – К.: Генеза, 2000. – 430 с.
4. Смирнов В.М., Дубровский В.И. Физиология физического воспитания и спорта. – М.: Физкультура и спорт, 2002. – 430 с.
5. Серопегин И.М., Волков В.М. Физиология человека. – М.: Просвещение, 1979. – 430 с.
6. Спортивная медицина. – М.: Медицина, 1975. – 370 с.
7. Теория и методика физического воспитания. – М.: Физкультура и спорт, 1976. – 280 с.
8. Яковлев Н.Я., Коробков А.В., Янанис С.В. Физиологические и биохимические основы теории и методики спортивной тренировки. – М.: Физкультура и спорт, 1960. – 317 с.
9. Теория спорта. – К.: Наука, 1987. – 380 с.
10. Уилмор Дж. Х., Костил Д.Н. Физиология спорта и двигательной активности. – К.: Наука, 1997 – 530 с.
11. Физиология мышечной деятельности. – М.: Медицина, 1982. – 310 с.
12. Холодов Ж.К., Кузнецов В.С. теория и методика физического воспитания и спорта. – М.: Академия, 2002. – 490 с.
13. Энциклопедия физической подготовки. – М.: Физкультура и спорт, 1994. – 560 с.
14. Васильева В., Коссовская Э., Степочкина Н. Физиология человека. – М.: Просвещение, 1973. – 330 с.
15. Проблемы физиологии спорта. – М.: Физкультура и спорт, 1960. – 390 с.
16. Вациорский В.М., Алешинский С.Ю., Якунин Н.А. Биохимические основы выносливости. – М.: Физкультура и спорт, 1982. – 395 с.