Эпителиальные ткани (от греч. epi - над и thele - кожица) - древнейшие гистологические структуры, которые в фило- и онтогенезе возникают первыми. Они представляют собой систему дифферонов полярно дифференцированных клеток, тесно расположенных в виде пласта на базальной мембране (пластинке), на границе с внешней или внутренней средой, а также образующих большинство желез организма. Различают поверхностные (покровные и выстилающие) и железистые эпителии.
6.1. ОБЩАЯ МОРФОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА И КЛАССИФИКАЦИИ
Поверхностные эпителии - это пограничные ткани, располагающиеся на поверхности тела (покровные), слизистых оболочек внутренних органов (желудка, кишечника, мочевого пузыря и др.) и вторичных полостей тела (выстилающие). Они отделяют организм и его органы от окружающей их среды и участвуют в обмене веществ между ними, осуществляя функции поглощения веществ (всасывание) и выделения продуктов обмена (экскреция). Например, через кишечный эпителий всасываются в кровь и лимфу продукты переваривания пищи, которые служат источником энергии и строительным материалом для организма, а через почечный эпителий выделяется ряд продуктов азотистого обмена, являющихся шлаками. Кроме этих функций, покровный эпителий выполняет важную защитную функцию, предохраняя подлежащие ткани организма от различных внешних воздействий - химических, механических, инфекционных и др. Например, кожный эпителий является мощным барьером для микроорганизмов и многих ядов. Наконец, эпителий, покрывающий внутренние органы, создает условия для их подвижности, например для сокращения сердца, экскурсии легких и т. д.
Железистый эпителий, образующий многие железы, осуществляет секреторную функцию, т. е. синтезирует и выделяет специфические продукты -
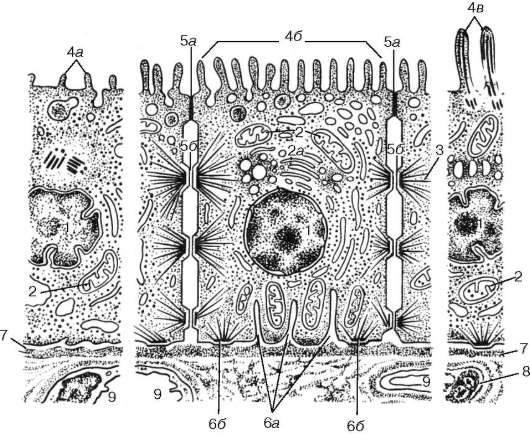
Рис. 6.1. Строение однослойного эпителия (по Е. Ф. Котовскому): 1 - ядро; 2 - митохондрии; 2а - комплекс Гольджи; 3 - тонофибриллы; 4 - структуры апикальной поверхности клеток: 4а - микроворсинки; 4б - микроворсинчатая (щеточная) каемка; 4в - реснички; 5 - структуры межклеточной поверхности: 5а - плотные контакты; 5б - десмосомы; 6 - структуры базальной поверхности клеток: 6а - инвагинации плазмолеммы; 6б - полудесмосомы; 7 - базальная мембрана (пластинка); 8 - соединительная ткань; 9 - кровеносные капилляры
секреты, которые используются в процессах, протекающих в организме. Например, секрет поджелудочной железы участвует в переваривании белков, жиров и углеводов в тонкой кишке, секреты эндокринных желез - гормоны - регулируют многие процессы (роста, обмена веществ и др.).
Эпителии участвуют в построении многих органов, в связи с чем обнаруживают большое разнообразие морфофизиологических свойств. Некоторые из них являются общими, позволяющими отличать эпителии от других тканей организма. Имеются следующие основные особенности эпителиев.
Эпителии представляют собой пласты клеток - эпителиоцитов (рис. 6.1), которые имеют неодинаковую форму и строение в различных видах эпителия. Между клетками, составляющими эпителиальный пласт, мало межклеточного вещества, и клетки тесно связаны друг с другом с помощью различных контактов - десмосом, промежуточных, щелевых и плотных соединений.
Эпителии располагаются на базальных мембранах, которые образуются в результате деятельности как клеток эпителия, так и подлежащей соединительной ткани. Базальная мембрана имеет толщину около 1 мкм и состоит из подэпителиальной электронно-прозрачной светлой пластинки

Рис. 6.2. Строение базальной мембраны (схема по Е. Ф. Котовскому): С - светлая пластинка (lamina lucida); T - темная пластинка (lamina densa); БМ - базальная мембрана. 1 - цитоплазма эпителиоцитов; 2 - ядро; 3 - прикрепительная пластинка полудесмосомы (гемидесмосомы); 4 - кератиновые тонофиламенты; 5 - якорные филаменты; 6 - плазмолемма эпителиоцитов; 7 - заякоривающие фибриллы; 8 - подэпителиальная рыхлая соединительная ткань; 9 - кровеносный капилляр
(lamina lucida) толщиной 20-40 нм и темной пластинки (lamina densa) толщиной 20-60 нм (рис. 6.2). Светлая пластинка включает аморфное вещество, относительно бедное белками, но богатое ионами кальция. Темная пластинка имеет богатый белками аморфный матрикс, в который впаяны фибриллярные структуры, обеспечивающие механическую прочность мембраны. В ее аморфном веществе содержатся сложные белки - гликопро-теины, протеогликаны и углеводы (полисахариды) - гликозаминогликаны. Гликопротеины - фибронектин и ламинин, выполняют функцию адгезивного субстрата, с помощью которого к мембране прикрепляются эпителио-циты. Важную роль при этом играют ионы кальция, обеспечивающие связь между адгезивными молекулами гликопротеинов базальной мембраны и полудесмосомами эпителиоцитов. Кроме того, гликопротеины индуцируют пролиферацию и дифференцировку эпителиоцитов при регенерации эпителия. Протеогликаны и гликозаминогликаны создают упругость мембраны и характерный для нее отрицательный заряд, от которого зависит ее избирательная проницаемость для веществ, а также способность накапливать в условиях патологии многие ядовитые вещества (токсины), сосудоактивные амины и комплексы из антигенов и антител.
Особенно прочно клетки эпителия связаны с базальной мембраной в области гемидесмосом (полудесмосом). Здесь от плазмолеммы базальных эпителиоцитов через светлую пластинку к темной пластинке базальной мембраны проходят «якор-
ные» филаменты. В этой же области, но со стороны подлежащей соединительной ткани в темную пластинку базальной мембраны вплетаются пучки «заякориваю-щих» фибрилл (содержат коллаген VII типа), обеспечивающих прочное прикрепление эпителиального пласта к подлежащей ткани.
Таким образом, базальная мембрана выполняет ряд функций: механическую (прикрепительную), трофическую и барьерную (избирательный транспорт веществ), морфогенетическую (организующую при регенерации) и ограничивающую возможность инвазивного роста эпителия.
В связи с тем, что в пласты эпителиоцитов не проникают кровеносные сосуды, питание эпителиоцитов осуществляется диффузно через базальную мембрану со стороны подлежащей соединительной ткани, с которой эпителий находится в тесном взаимодействии.
Эпителий обладает полярностью, т. е. базальные и апикальные отделы эпителиоцитов имеют разное строение. В однослойных эпителиях наиболее отчетливо выражена полярность клеток, проявляющаяся морфологическими и функциональными различиями апикальной и базальной частей эпите-лиоцитов. Так, эпителиоциты тонкой кишки имеют на апикальной поверхности множество микроворсинок, обеспечивающих всасывание продуктов пищеварения. В базальной части эпителиоцита микроворсинки отсутствуют, через нее осуществляются всасывание и выделение в кровь или лимфу продуктов обмена. В многослойных эпителиях, кроме того, отмечается полярность пласта клеток - различие в строении эпителиоцитов базально-го, промежуточного и поверхностного слоев (см. рис. 6.1).
Эпителиальные ткани, как правило, относятся к обновляющимся тканям. Поэтому им присуща высокая способность к регенерации. Восстановление эпителия происходит вследствие митотического деления и дифференци-ровки камбиальных клеток. В зависимости от места расположения камбиальных клеток в эпителиальных тканях различают диффузный и локализованный камбий.
Источники развития и классификации эпителиальных тканей. Эпителии развиваются из всех трех зародышевых листков, начиная с 3-4-й недели эмбрионального развития человека. В зависимости от эмбрионального источника различают эпителии эктодермального, мезодермального и энто-дермального происхождения. Эпителиоциты формируют клеточные пласты и являются ведущим клеточным диффероном в данной ткани. В гистогенезе в состав эпителия (кроме эпителиоцитов) могут входить гистологические элементы дифферонов иного происхождения (сопутствующие диффероны в полидифферонных эпителиях). Существуют также эпителии, где наряду с пограничными эпителиоцитами, в результате дивергентной дифферен-цировки стволовой клетки возникают клеточные диффероны эпителиоци-тов секреторной и эндокринной специализации, интегрированные в состав эпителиального пласта. Лишь родственные виды эпителиев, развивающиеся из одного зародышевого листка, в условиях патологии могут подвергаться метаплазии, т. е. переходить из одного вида в другой, например, в дыхательных путях эктодермальный эпителий при хронических бронхитах из однослойного реснитчатого может превратиться в многослойный плоский,
который в норме характерен для полости рта и имеет также эктодермаль-ное происхождение.
Цитохимическим маркёром эпителиоцитов является белок цитокера-тин, образующий промежуточные филаменты. В разных видах эпителиев он имеет различные молекулярные формы. Известно более 20 форм этого белка. Иммуногистохимическое выявление этих форм цитокератина позволяет определить принадлежность исследуемого материала к тому или иному типу эпителиев, что имеет большое значение в диагностике опухолей.
Классификации. Существует несколько классификаций эпителиев, в основу которых положены различные признаки: происхождение, строение, функция. При построении классификаций учитываются гистологические признаки, характеризующие ведущий клеточный дифферон. Наибольшее распространение получила морфологическая классификация, учитывающая главным образом отношение клеток к базальной мембране и их форму (схема 6.1).
Согласно этой классификации, среди покровных и выстилающих эпите-лиев, входящих в состав кожи, серозных и слизистых оболочек внутренних органов (полость рта, пищевод, пищеварительный тракт, органы дыхания, матка, мочеотводящие пути и др.) различают две основные группы эпите-лиев: однослойные и многослойные. В однослойных эпителиях все клетки связаны с базальной мембраной, а в многослойных с ней непосредственно связан лишь один нижний слой клеток, а остальные вышележащие слои такой связи не имеют. В соответствии с формой клеток, составляющих однослойные эпителии, последние подразделяются на плоские (сквамозные), кубические и столбчатые (призматические). В определении многослойных эпителиев учитывается лишь форма клеток наружных слоев. Например, эпителий роговицы глаза - многослойный плоский, хотя его нижние слои состоят из клеток столбчатой и крылатой формы.
Однослойный эпителий может быть однорядным и многорядным. У однорядного эпителия все клетки имеют одинаковую форму - плоскую, кубическую или столбчатую, их ядра расположены на одном уровне, т. е. в один ряд. Такой эпителий называют еще изоморфным (от греч. isos - равный). Однослойный эпителий, имеющий клетки различной формы и высоты, ядра которых лежат на разных уровнях, т. е. в несколько рядов, носит название многорядного, или псевдомногослойного (анизоморфного).
Многослойный эпителий бывает ороговевающим, неороговевающим и переходным. Эпителий, в котором протекают процессы ороговения, связанные с дифференцировкой клеток верхних слоев в плоские роговые чешуйки, называют многослойным плоским ороговевающим. При отсутствии ороговения эпителий является многослойным плоским неороговевающим.
Переходный эпителий выстилает органы, подверженные сильному растяжению, - мочевой пузырь, мочеточники и др. При изменении объема органа толщина и строение эпителия также изменяются.
Наряду с морфологической классификацией используется онтофилогене-тическая классификация, созданная российским гистологом Н. Г. Хлопиным. В зависимости от эмбрионального зачатка, служащего источником развития

Схема 6.1. Морфологическая классификация видов поверхностного эпителия
ведущего клеточного дифферона, эпителии подразделяются на типы: эпи-дермальный (кожный), энтеродермальный (кишечный), целонефродермаль-ный, эпендимоглиальный и ангиодермальный типы эпителиев.
Эпидермальный тип эпителия образуется из эктодермы, имеет многослойное или многорядное строение, приспособлен к выполнению прежде всего защитной функции (например, многослойный плоский ороговеваю-щий эпителий кожи).
Энтеродермальный тип эпителия развивается из энтодермы, является по строению однослойным призматическим, осуществляет процессы всасывания веществ (например, однослойный каемчатый эпителий тонкой кишки), выполняет железистую функцию (например, однослойный эпителий желудка).
Целонефродермальный тип эпителия развивается из мезодермы, по строению однослойный, плоский, кубический или призматический; выполняет главным образом барьерную или экскреторную функцию (например, плоский эпителий серозных оболочек - мезотелий, кубический и призматический эпителии в мочевых канальцах почек).
Эпендимоглиальный тип представлен специальным эпителием, выстилающим, например, полости мозга. Источником его образования является нервная трубка.
К ангиодермальному типу эпителия относят эндотелиальную выстилку кровеносных сосудов. По строению эндотелий подобен однослойным плоским эпителиям. Его принадлежность к эпителиальным тканям являет-
ся спорной. Многие исследователи относят эндотелий к соединительной ткани, с которой он связан общим эмбриональным источником развития - мезенхимой.
6.1.1. Однослойные эпителии
Однорядные эпителии
Однослойный плоский эпителий (epithelium simplex squamosum) представлен в организме мезотелием и, по некоторым данным, эндотелием.
Мезотелий (mesothelium) покрывает серозные оболочки (листки плевры, висцеральную и париетальную брюшину, околосердечную сумку). Клетки мезотелия - мезотелиоциты - плоские, имеют полигональную форму и неровные края (рис. 6.3, а). В той части, где в них располагается ядро, клетки более «толстые». Некоторые из них содержат не одно, а два или даже три ядра, т. е. полиплоидные. На свободной поверхности клетки имеются микроворсинки. Через мезотелий происходят выделение и всасывание серозной жидкости. Благодаря его гладкой поверхности легко осуществляется скольжение внутренних органов. Мезотелий препятствует образованию соединительнотканных спаек между органами брюшной и грудной полостей, развитие которых возможно при нарушении его целости. Среди мезотелиоцитов присутствуют малодифференцированные (камбиальные) формы, способные к размножению.
Эндотелий (endothelium) выстилает кровеносные и лимфатические сосуды, а также камеры сердца. Он представляет собой пласт плоских клеток - эндоте-лиоцитов, лежащих в один слой на базальной мембране. Эндотелиоциты относительно бедны органеллами, в их цитоплазме присутствуют пиноцитозные пузырьки. Эндотелий, располагаясь в сосудах на границе с лимфой, кровью, участвует в обмене веществ и газов (О2, СО2) между ними и другими тканями. Эндотелиоциты синтезируют разнообразные факторы роста, вазоактивные вещества и др. При повреждении эндотелия возможны изменение кровотока в сосудах и образование в их просвете сгустков крови - тромбов. В различных участках сосудистой системы эндотелиоциты различаются размерами, формой и ориентацией относительно оси сосуда. Эти свойства эндотелиоцитов обозначаются как гетероморфия, или полиморфия (Н. А. Шевченко). Эндотелиоциты, способные к размножению, располагаются диффузно, с преобладанием в зонах дихотомического деления сосуда.
Однослойный кубический эпителий (epithelium simplex cuboideum) выстилает часть почечных канальцев (проксимальные и дистальные). Клетки проксимальных канальцев имеют микроворсинчатую (щеточную) каемку и базальную исчерченность. Щеточная каемка состоит из большого числа микроворсинок. Исчерченность обусловлена наличием в базальных отделах клеток глубоких складок плазмолеммы и митохондриями, расположенными между ними. Эпителий почечных канальцев выполняет функцию обратного всасывания (реабсорбция) ряда веществ из первичной мочи, протекающей по канальцам, в кровь межканальцевых сосудов. Камбиальные клетки

Рис. 6.3. Строение однослойных эпителиев:
а - плоский эпителий (мезотелий); б - столбчатый микроворсинчатый эпителий: 1 - микроворсинки (каемка); 2 - ядро эпителиоцита; 3 - базальная мембрана; 4 - соединительная ткань; в - микрофотография: 1 - каемка; 2 - микроворсинчатые эпителиоциты; 3 - бокаловидная клетка; 4 - соединительная ткань
располагаются диффузно среди эпителиоцитов. Однако пролиферативная активность клеток крайне низкая.
Однослойный столбчатый (призматический) эпителий (epithelium simplex columnare). Этот вид эпителия характерен для среднего отдела пищеварительной системы (см. рис. 6.3, б, в). Он выстилает внутреннюю поверхность желудка, тонкой и толстой кишки, желчного пузыря, ряда протоков печени и поджелудочной железы. Эпителиальные клетки связаны между собой с помощью десмосом, щелевых коммуникационных соединений, по типу замка, плотных замыкающих соединений (см. главу 4). Благодаря последним в межклеточные щели эпителия не может проникнуть содержимое полости желудка, кишки и других полых органов.
В желудке в однослойном столбчатом эпителии все клетки являются железистыми (поверхностными мукоцитами), продуцирующими слизь. Секрет мукоцитов защищает стенку желудка от грубого влияния комков пищи и переваривающего действия желудочного сока, имеющего кислую реакцию, и ферментов, расщепляющих белки. Меньшая часть клеток эпителия, расположенных в желудочных ямочках - мелких углублениях в стенке желудка, представляет собой камбиальные эпителио-циты, способные делиться и дифференцироваться в железистые эпителиоциты. За счет ямочных клеток каждые 5 сут происходит полное обновление эпителия желудка - его физиологическая регенерация.
В тонкой кишке эпителий однослойный столбчатый, активно участвующий в пищеварении, т. е. в расщеплении пищи до конечных продуктов и всасывании их в кровь и лимфу. Он покрывает в кишке поверхность ворсинок и образует стенку кишечных желез - крипт. Эпителий ворсинок в основном состоит из микроворсинчатых эпителиоцитов. Микроворсинки апикальной поверхности эпителиоцита покрыты гли-кокаликсом. Здесь происходит мембранное пищеварение - расщепление (гидролиз) веществ пищи до конечных продуктов и всасывание их (транспорт через мембрану и цитоплазму эпителиоцитов) в кровеносные и лимфатические капилляры подлежащей соединительной ткани. В той части эпителия, который выстилает крипты кишки, различают бескаемчатые столбчатые эпителиоциты, бокаловидные клетки, а также эндокринные клетки и экзокриноциты с ацидофильными гранулами (клетки Панета). Бескаемчатые эпителиоциты крипт являются камбиальными клетками кишечного эпителия, способными к пролиферации (размножению) и дивергентной дифференци-ровке в микроворсинчатые, бокаловидные, эндокринные и в клетки Панета. Благодаря камбиальным клеткам микроворсинчатые эпителиоциты полностью обновляются (регенерируют) в течение 5-6 сут. Бокаловидные клетки выделяют слизь на поверхность эпителия. Слизь защищает его и подлежащие ткани от механических, химических и инфекционных воздействий, а также участвует в пристеночном пищеварении, т. е. в расщеплении белков, жиров и углеводов пищи с помощью адсорбированных в ней ферментов до промежуточных продуктов. Эндокринные (базально-зернистые) клетки нескольких видов (ЕС, D, S и др.) секретируют в кровь гормоны, которые осуществляют местную регуляцию функции органов пищеварительного аппарата. Клетки Панета вырабатывают лизоцим - бактерицидное вещество.
Однослойные эпителии также представлены производными нейроэктодер-мы - эпителии эпендимоглиального типа. По строению клеток он варьирует от плоского до столбчатого. Так, эпендимный эпителий, выстилающий центральный канал спинного мозга и желудочки головного мозга, является однослойным столбчатым. Пигментный эпителий сетчатки глаза представляет собой однослойный эпителий, состоящий из клеток полигональной формы. Периневральный эпителий, окружающий нервные стволы и выстилающий периневральное пространство, - однослойный плоский. Как производные нейроэктодермы, эпителии обладают ограниченными возможностями регенерации, преимущественно внутриклеточным способом.
Многорядные эпителии
Многорядные (псевдомногослойные) эпителии (epithelium pseudostrati-ficatum) выстилают воздухоносные пути - носовую полость, трахею, бронхи, а также ряд других органов. В воздухоносных путях многорядный столбчатый эпителий является реснитчатым. Разнообразие видов клеток

Рис. 6.4. Строение многорядного столбчатого реснитчатого эпителия: а - схема: 1 - мерцательные реснички; 2 - бокаловидные клетки; 3 - реснитчатые клетки; 4 - вставочные клетки; 5 - базальные клетки; 6 - базальная мембрана; 7 - соединительная ткань; б - микрофотография: 1 - реснички; 2 - ядра реснитчатых и вставочных клеток; 3 - базальные клетки; 4 - бокаловидные клетки; 5 - соединительная ткань
в составе эпителия (реснитчатые, вставочные, базальные, бокаловидные, клетки Клара и эндокринные клетки) есть результат дивергентной диффе-ренцировки камбиальных (базальных) эпителиоцитов (рис. 6.4).
Базальные эпителиоциты низкие, располагаются на базальной мембране в глубине эпителиального пласта, участвуют в регенерации эпителия. Реснитчатые (мерцательные) эпителиоциты высокие, столбчатой (призматической) формы. Эти клетки составляют ведущий клеточный дифферон. Их апикальная поверхность покрыта ресничками. Движение ресничек обеспечивает транспорт слизи и инородных частиц по направлению к глотке (мукоцилиарный транспорт). Бокаловидные эпителиоциты секретируют на поверхность эпителия слизь (муцины), которая защищает его от механических, инфекционных и других воздействий. В эпителии также присутствуют несколько видов эндокриноцитов (ЕС, D, Р), гормоны которых осуществляют местную регуляцию мышечной ткани воздухоносных путей. Все эти виды клеток имеют разную форму и размеры, поэтому их ядра располагаются на разных уровнях эпителиального пласта: в верхнем ряду - ядра реснитчатых клеток, в нижнем - ядра базальных клеток, а в среднем - ядра вставочных, бокаловидных и эндокринных клеток. Кроме эпителиальных дифферонов, в составе многорядного столбчатого эпителия присутствуют гистологические элементы гематогенного дифферона (специализированные макрофаги, лимфоциты).
6.1.2. Многослойные эпителии
Многослойный плоский неороговевающий эпителий (epithelium stiatificatum squamosum noncornificatum) покрывает снаружи роговицу глаза, выстилает

Рис. 6.5. Строение многослойного плоского неороговевающего эпителия роговицы глаза (микрофотография): 1 - слой плоских клеток; 2 - шиповатый слой; 3 - базальный слой; 4 - базальная мембрана; 5 - соединительная ткань
полости рта и пищевода. В нем различают три слоя: базальный, шиповатый (промежуточный) и поверхностный (рис. 6.5). Базальный слой состоит из эпителиоцитов столбчатой формы, располагающихся на базальной мембране. Среди них имеются камбиальные клетки, способные к митотическому делению. За счет вновь образованных клеток, вступающих в дифференциров-ку, происходит смена эпителиоци-тов вышележащих слоев эпителия. Шиповатый слой состоит из клеток неправильной многоугольной формы. В эпителиоцитах базально-го и шиповатого слоев хорошо развиты тонофибриллы (пучки тоно-филаментов из белка кератина), а между эпителиоцитами - десмосомы и другие виды контактов. Поверхностные слои эпителия образованы плоскими клетками. Заканчивая свой жизненный цикл, последние отмирают и отпадают.
Многослойный плоский ороговевающий эпителий (epithelium stratificatum squamosum comificatum) (рис. 6.6) покрывает поверхность кожи, образуя ее эпидермис, в котором происходит процесс ороговения (кератинизации), связанный с дифференцировкой эпителиальных клеток - кератиноцитов в роговые чешуйки наружного слоя эпидермиса. Дифференцировка керати-ноцитов проявляется их структурными изменениями в связи с синтезом и накоплением в цитоплазме специфических белков - цитокератинов (кислых и щелочных), филаггрина, кератолинина и др. В эпидермисе различают несколько слоев клеток: базальный, шиповатый, зернистый, блестящий и роговой. Последние три слоя особенно сильно выражены в коже ладоней и подошв.
Ведущий клеточный дифферон в эпидермисе представлен кератиноцита-ми, которые по мере дифференцировки перемещаются из базального слоя в вышележащие слои. Кроме кератиноцитов, в эпидермисе находятся гистологические элементы сопутствующих клеточных дифферонов - меланоциты (пигментные клетки), внутриэпидермальные макрофаги (клетки Лангерганса), лимфоциты и клетки Меркеля.
Базальный слой состоит из столбчатых по форме кератиноцитов, в цитоплазме которых синтезируется кератиновый белок, формирующий тоно-филаменты. Здесь же находятся камбиальные клетки дифферона кератино-цитов. Шиповатый слой образован кератиноцитами многоугольной формы, которые прочно связаны между собой многочисленными десмосомами. В месте десмосом на поверхности клеток имеются мельчайшие выросты -
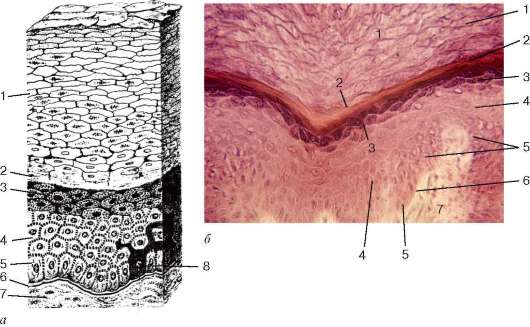
Рис. 6.6. Многослойный плоский ороговевающий эпителий:
а - схема: 1 - роговой слой; 2 - блестящий слой; 3 - зернистый слой; 4 - шиповатый слой; 5 - базальный слой; 6 - базальная мембрана; 7 - соединительная ткань; 8 - пигментоцит; б - микрофотография
«шипики», у смежных клеток направленные навстречу друг другу. Они хорошо заметны при расширении межклеточных пространств или при сморщивании клеток, а также при мацерации. В цитоплазме шиповатых кератиноцитов тонофиламенты образуют пучки - тонофибриллы и появляются кератиносомы - гранулы, содержащие липиды. Эти гранулы путем экзоцитоза выделяются в межклеточное пространство, где образуют богатое липидами вещество, цементирующее кератиноциты.
В базальном и шиповатом слоях также присутствуют отростчатой формы меланоциты с гранулами черного пигмента - меланина, клетки Лангерганса (дендритные клетки) и клетки Меркеля (тактильные эпителиоциты), имеющие мелкие гранулы и контактирующие с афферентными нервными волокнами (рис. 6.7). Меланоциты с помощью пигмента создают барьер, препятствующий проникновению в организм ультрафиолетовых лучей. Клетки Лангерганса являются разновидностью макрофагов, участвуют в защитных иммунных реакциях и регулируют размножение (деление) кератиноцитов, образуя вместе с ними «эпидермально-пролиферативные единицы». Клетки Меркеля являются чувствительными (осязательными) и эндокринными (апудоцитами), влияющими на регенерацию эпидермиса (см. главу 15).
Зернистый слой состоит из уплощенных кератиноцитов, в цитоплазме которых содержатся крупные базофильные гранулы, получившие название кератогиалиновых. Они включают промежуточные филаменты (кератин) и синтезируемый в кератиноцитах этого слоя белок - филаггрин, а

Рис. 6.7. Строение и клеточно-дифферонный состав многослойного плоского орого-вевающего эпителия (эпидермиса) (по Е. Ф. Котовскому):
I - базальный слой; II - шиповатый слой; III - зернистый слой; IV, V - блестящий и роговой слои. К - кератиноциты; Р - корнеоциты (роговые чешуйки); М - макрофаг (клетка Лангерганса); Л - лимфоцит; О - клетка Меркеля; П - меланоцит; С - стволовая клетка. 1 - митотически делящийся кератиноцит; 2 - кератиновые тонофиламенты; 3 - десмосомы; 4 - кератиносомы; 5 - кератогиалиновые гранулы; 6 - слой кератолинина; 7 - ядро; 8 - межклеточное вещество; 9, 10 - керати-новые фибриллы; 11 - цементирующее межклеточное вещество; 12 - отпадающая чешуйка; 13 - гранулы в форме теннисных ракеток; 14 - базальная мембрана; 15 - сосочковый слой дермы; 16 - гемокапилляр; 17 - нервное волокно
также вещества, образующиеся в результате начинающегося здесь распада органелл и ядер под влиянием гидролитических ферментов. Кроме того, в зернистых кератиноцитах синтезируется еще один специфический белок - кератолинин, укрепляющий плазмолемму клеток.
Блестящий слой выявляется только в сильно ороговевающих участках эпидермиса (на ладонях и подошвах). Он образован постклеточными структурами. В них отсутствуют ядра и органеллы. Под плазмолеммой располагается электронно-плотный слой из белка кератолинина, придающего ей прочность и защищающего от разрушительного действия гидролитических ферментов. Кератогиалиновые гранулы сливаются, и внутренняя часть клеток заполняется светопреломляющей массой из кератиновых фибрилл, склеенных аморфным матриксом, содержащим филаггрин.
Роговой слой очень мощный в коже пальцев, ладоней, подошв и относительно тонкий в остальных участках кожи. Он состоит из плоских многоугольной формы (тетрадекаэдр) роговых чешуек, имеющих толстую оболочку с кератолинином и заполненных кератиновыми фибриллами, расположенными в аморфном матриксе, состоящем из другого вида кератина. Филаггрин при этом распадается на аминокислоты, которые входят в состав кератина фибрилл. Между чешуйками находится цементирующее вещество - продукт кератиносом, богатый липидами (церамидами и др.) и поэтому обладающий гидроизолирующим свойством. Самые наружные роговые чешуйки утрачивают связь друг с другом и постоянно отпадают с поверхности эпителия. На смену им приходят новые - вследствие размножения, дифференцировки и перемещения клеток из нижележащих слоев. Благодаря этим процессам, составляющим физиологическую регенерацию, в эпидермисе полностью обновляется состав кератиноцитов через каждые 3-4 нед. Значение процесса кератинизации (ороговения) в эпидермисе заключается в том, что образующийся при этом роговой слой обладает устойчивостью к механическим и химическим воздействиям, плохой тепло-проводимостью и непроницаемостью для воды и многих водорастворимых ядовитых веществ.
Переходный эпителий (epithelium transitionale). Этот вид многослойного эпителия типичен для мочеотводящих органов - лоханок почек, мочеточников, мочевого пузыря, стенки которых подвержены значительному растяжению при заполнении мочой. В нем различают несколько слоев клеток - базальный, промежуточный, поверхностный (рис. 6.8, а, б).

Рис. 6.8. Строение переходного эпителия (схема):
а - при нерастянутой стенке органа; б - при растянутой стенке органа. 1 - переходный эпителий; 2 - соединительная ткань
Базальный слой образован мелкими почти округлыми (темными) камбиальными клетками. В промежуточном слое располагаются клетки полигональной формы. Поверхностный слой состоит из очень крупных, нередко дву- и трехъядерных клеток, имеющих куполообразную или уплощенную форму в зависимости от состояния стенки органа. При растяжении стенки вследствие заполнения органа мочой эпителий становится более тонким и его поверхностные клетки уплощаются. Во время сокращения стенки органа толщина эпителиального пласта резко возрастает. При этом некоторые клетки в промежуточном слое «выдавливаются» кверху и принимают грушевидную форму, расположенные над ними поверхностные клетки - куполообразную форму. Между поверхностными клетками обнаружены плотные контакты, имеющие значение для предотвращения проникновения жидкости через стенку органа (например, мочевого пузыря).
Регенерация. Покровный эпителий, занимая пограничное положение, постоянно испытывает влияние внешней среды, поэтому эпителиальные клетки сравнительно быстро изнашиваются и погибают. Источником их восстановления являются камбиальные клетки эпителия, которые обеспечивают клеточную форму регенерации, так как сохраняют способность к делению в течение всей жизни организма. Размножаясь, часть вновь образованных клеток вступают в дифференцировку и превращаются в эпителиоциты, подобные утраченным. Камбиальные клетки в многослойных эпителиях находятся в базальном (зачатковом) слое, в многорядных эпителиях к ним относятся базальные клетки, в однослойных эпителиях они располагаются в определенных участках: например, в тонкой кишке - в эпителии крипт, в желудке - в эпителии ямочек, а также шеек собственных желез, в мезотелии - среди мезотелиоцитов и т. п. Высокая способность большинства эпителиев к физиологической регенерации служит основой для его быстрого восстановления в патологических условиях (репаративная регенерация). Напротив, производные нейроэктодермы восстанавливаются преимущественно внутриклеточным способом.
С возрастом в покровном эпителии наблюдается ослабление процессов обновления клеток.
Иннервация. Эпителий хорошо иннервирован. В нем имеются многочисленные чувствительные нервные окончания - рецепторы.
6.2. ЖЕЛЕЗИСТЫЕ ЭПИТЕЛИИ
Для этих эпителиев характерна секреторная функция. Железистый эпителий (epithelium glandulare) состоит из железистых, или секреторных, эпи-телиоцитов (гландулоцитов). Они осуществляют синтез, а также выделение специфических продуктов - секретов на поверхность кожи, слизистых оболочек и в полости ряда внутренних органов (внешняя - экзокринная секреция) или в кровь и лимфу (внутренняя - эндокринная секреция).
Путем секреции в организме выполняются многие важные функции: образование молока, слюны, желудочного и кишечного сока, желчи, эндо-
кринная (гуморальная) регуляция и др. Большинство клеток отличаются наличием секреторных включений в цитоплазме, хорошо развитыми эндо-плазматической сетью и комплексом Гольджи, полярным расположением органелл и секреторных гранул.
Секреторные эпителиоциты лежат на базальной мембране. Форма их весьма разнообразна и меняется в зависимости от фазы секреции. Ядра бывают обычно крупными, часто неправильной формы. В цитоплазме клеток, которые вырабатывают секреты белкового характера (например, пищеварительные ферменты), хорошо развита гранулярная эндоплазматическая сеть. В клетках, синтезирующих небелковые секреты (липиды, стероиды), выражена агранулярная эндоплазматическая сеть. Комплекс Гольджи обширный. Его форма и расположение в клетке меняются в зависимости от фазы секреторного процесса. Митохондрии, как правило, многочисленны. Они накапливаются в местах наибольшей активности клеток, т. е. там, где образуется секрет. В цитоплазме клеток обычно присутствуют секреторные гранулы, размер и строение которых зависят от химического состава секрета. Число их колеблется в связи с фазами секреторного процесса. В цитоплазме некоторых гландулоцитов (например, участвующих в образовании соляной кислоты в желудке) обнаруживаются внутриклеточные секреторные канальцы - глубокие впячивания плазмолеммы, покрытые микроворсинками. Плазмолемма имеет различное строение на латеральных, базальных и апикальных поверхностях клеток. На первых она образует десмосомы и плотные запирающие контакты. Последние окружают верхушечные (апикальные) части клеток, отделяя таким образом межклеточные щели от просвета железы. На базальных поверхностях клеток плазмолемма образует небольшое число узких складок, проникающих в цитоплазму. Такие складки особенно хорошо развиты в клетках желез, выделяющих секрет, богатый солями, например в клетках выводных протоков слюнных желез. Апикальная поверхность клеток покрыта микроворсинками.
В железистых клетках хорошо заметна полярная дифференцировка. Она обусловлена направленностью секреторных процессов, например при внешней секреции от базальной к апикальной части клетки.
Периодические изменения железистой клетки, связанные с образованием, накоплением, выделением секрета и восстановлением ее для дальнейшей секреции, получили название секреторного цикла.
Для образования секрета из крови и лимфы в железистые клетки со стороны б