Задняя теменная кора
Материал из Википедии — свободной энциклопедии
Переход к навигацииПереход к поиску
| Задняя теменная кора | |
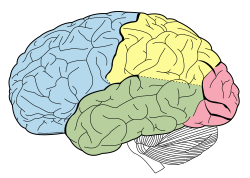 Доли головного мозга. Теменная доля желтая, а задняя часть находится рядом с красной областью.
Доли головного мозга. Теменная доля желтая, а задняя часть находится рядом с красной областью.
| |
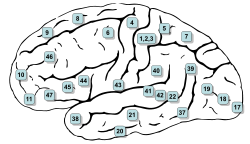 Боковая поверхность мозга с пронумерованными областями Бродмана. (№5 и №7 в правом верхнем углу)
Боковая поверхность мозга с пронумерованными областями Бродмана. (№5 и №7 в правом верхнем углу)
| |
| Подробные сведения | |
| Идентификаторы | |
| Латинский | Задняя теменная кора головного мозга |
| Анатомические термины нейроанатомии [редактировать в Викиданных] |
Задняя теменная кора (часть теменной неокортекса, расположенная за первичной соматосенсорной корой) играет важную роль в планируемых движениях, пространственном мышлении и внимании.
Повреждение задней теменной коры может вызвать различные сенсомоторные нарушения, в том числе нарушения восприятия и памяти пространственных отношений, неточное достижение и схватывание, контроль за движением глаз и невнимательность. Двумя наиболее поразительными последствиями повреждения КПП являются апраксия и пренебрежение пространством[1].
Содержание
- 1Анатомия
- 2Функции
- 2.1Двигатель
- 2.2Другое
- 3Рекомендации
- 4Внешние ссылки
Анатомия[править]
Задняя теменная кора получает информацию от трех сенсорных систем, которые играют роль в локализации тела и внешних объектов в пространстве: зрительной системы, слуховой системы и соматосенсорной системы. В свою очередь, большая часть продукции задней теменной коры направляется в области лобной моторной коры: дорсолатеральную префронтальную кору, различные области вторичной моторной коры и фронтальное поле зрения.
Задняя теменная кора делится на внутритеменной борозды, чтобы сформировать спинной улучшенный теменной дольки и брюшной уступает теменной дольки.[2][3][4] зона Бродмана 7-это часть верхней теменной дольки,[2][5], но некоторые источники включают зона Бродмана 5.[5] нижней теменной дольки далее подразделяются на запредельного извилиныв теменной стыки угловой извилины.[2][3][4] нижней теменной дольки соответствует зоны Бродмана 39 и 40.[2][4]
Функции[править]
Двигатель[править]
Считается, что задняя теменная кора имеет отдельные представления для различных моторных эффекторов (например, рука против глаза).[6]
В дополнение к разделению по типу эффектора, некоторые области активируются как во время принятия решения, так и во время выполнения, в то время как другие области активны только во время выполнения. В одном исследовании записи одной клетки показали активность в области теменной досягаемости, в то время как приматы, не являющиеся людьми, решали, достигать цели или совершать саккаду, и активность сохранялась во время выбранного движения, если и только если обезьяна решила сделать движение для достижения цели. Однако клетки в области 5d были активны только после того, как было принято решение дотянуться рукой.[7] Другое исследование показало, что нейроны в области 5d кодируют только следующее движение в последовательности движений достижения, а не движения достижения позже в последовательности.[8]
В другом эксперименте с записью одной клетки нейроны в области теменной досягаемости демонстрировали реакции, соответствующие любому из двух целевых местоположений в последовательности запланированных движений достижения, предполагая, что различные части запланированной последовательности местоположений могут быть представлены параллельно в области теменной досягаемости.[9]
Задняя теменная кора, по-видимому, участвует в обучении двигательным навыкам. В исследовании с домашними животными исследователи попросили испытуемых научиться водить рукой по лабиринту. Активация в правой задней теменной коре наблюдалась во время выполнения задания, и снижение активации было связано с количеством допущенных ошибок.[10] Изучение интерфейса мозг-компьютер приводит к аналогичной схеме: активация задней теменной коры уменьшалась по мере того, как испытуемые становились более опытными.[11] Одно исследование показало, что начинающие художники увеличивают кровоток в правой задней теменной области по сравнению с художниками-экспертами, когда им приходится решать задачи, связанные с искусством.[12]
В исследовании, проведенном нейробиологами из Нью-Йоркского университета, когерентные паттерны срабатывания нейронов в КПП головного мозга были связаны с координацией различных эффекторов. Исследователи изучили неврологическую активность макак, заставляя их выполнять различные задачи, которые требовали от них либо достигать и одновременно использовать быстрые движения глаз (саккады), либо использовать только саккады. Когерентный паттерн срабатывания нейронов в КПП был виден только тогда, когда и глаза, и руки требовались для выполнения одной и той же задачи, но не для задач, которые включали только саккады.[13]
Кроме того, нейроны в задней теменной коре одновременно кодируют различные аспекты планируемого действия. Куанг и его коллеги обнаружили, что нейроны PPC кодируют не только запланированное физическое движение, но и ожидаемые визуальные последствия запланированного движения в течение периода планирования.[14]
Другое[править]
Исследования показывают, что височно-теменное соединение участвует в экзогенном или стимулируемом внимании, в то время как верхняя теменная долька демонстрирует временную активацию для самонаправленных переключений внимания.[15] Поддержание пространственного внимания зависит от правой задней теменной коры; повреждения в области между внутрипариетальной бороздой и нижней теменной долькой в правом ППК были значительно связаны с дефицитом устойчивого пространственного внимания.[16]
Задняя теменная кора последовательно активируется во время эпизодического извлечения, но большинство гипотез о том, почему это происходит, являются умозрительными и обычно устанавливают некоторую связь между вниманием и эпизодическим воспоминанием.[2][3]
Повреждение задней теменной коры приводит к дефициту зрительной рабочей памяти.[17] Пациенты могли называть объекты, которые они видели ранее, но были нарушены при распознавании ранее представленных объектов, даже если эти объекты имели знакомое название.
В другой парадигме рабочей памяти участники должны были по-разному реагировать на одни и те же стимулы (буквы X/Y) на основе предыдущих стимулов.[18] Предыдущие стимулы состояли из контекста более низкого уровня (буквы A/B) и контекста более высокого уровня (числа 1/2). Нижний контекст определял соответствующие реакции на стимулы X/Y, в то время как контекст более высокого уровня сигнализировал об изменении эффекта контекста более низкого уровня. Задняя теменная кора была активирована обновлениями контекста более низкого уровня, но не обновлениями контекста более высокого уровня.
Задняя теменная кора также активируется во время задач по рассуждению, и некоторые области, активированные для рассуждения, как правило, также активируются для математики или вычислений.[19]
Существуют также доказательства, указывающие на то, что он играет определенную роль в восприятии боли[20].
Недавние исследования показали, что чувство "свободы воли", по крайней мере частично, возникает в этой области.[21][22]
Рекомендации[править]
1. ^ Пинель, Джон П. Дж.Биопсихология Седьмое издание. Pearson Education Inc., 2009
2. ^ Перейти к:a b c d e Кабеса Р., Чиарамелли Э., Олсон И. Р., Москович М. (2008). "Теменная кора и эпизодическая память: отчет о внимании". Природа Рассматривает Неврологию. 9 (8): 613–625. doi:10.1038/nrn2459. ЧВК 2692883. PMID 18641668.
3. ^ Перейти к:a b c Хатчинсон Дж. Б., Ункафер М. Р., Вагнер А. Д. (2009). "Задняя теменная кора и эпизодическое извлечение: конвергентные и дивергентные эффекты внимания и памяти". Обучение и память. 16 (6): 343–356. doi:10.1101/lm.919109. ЧВК 2704099. PMID 19470649.
4. ^ Подпрыгните до:a b c Мартин, Р. Е. (н. д.). Давайте познакомимся с теменными долями! [PDF]. Извлечено из https://gablab.mit.edu/downloads/Parietal_Primer.pdf Архивировано 2018-02-18 на машине обратного пути
5. ^ Перейти к:a b Scheperjans F., Hermann K., Eickhoff S. B., Amunts K., Schleicher A., Zilles K. (2007). "Независимое от наблюдателя цитоархитектоническое картирование верхней теменной коры головного мозга человека". Кора головного мозга. 18 (4): 846–867. doi:10.1093/cercor/bhm116. PMID 17644831.
6. ^ Hwang E., Hauschild M., Wilke M., Andersen R. (2012). "Инактивация области теменной досягаемости Вызывает оптическую атаксию, Ухудшая Досягаемость, но не Саккады". Нейрон. 76 (5): 1021-1029. doi:10.1016/j.нейрон.2012.10.030. ЧВК 3597097. PMID 23217749.
7. ^ Цуй Х., Андерсен Р. А. (2011). "Различные представления Потенциальных и выбранных двигательных планов различными теменными областями". Журнал неврологии. 31 (49): 18130-18136. doi:10.1523/jneurosci.6247-10.2011. PMC 3327481. PMID 22159124.
8. ^ Ли Ю., Цуй Х. (2013). "Дорсальная Теменная область 5 Кодирует Немедленную Досягаемость Последовательными Движениями Рук". Журнал неврологии. 33 (36): 14455-14465. doi:10.1523/jneurosci.1162-13.2013. ЧВК 6618382. PMID 24005297.
9. ^ Baldauf D., Cui H., Andersen R. A. (2008). "Задняя теменная кора Параллельно Кодирует Обе Цели для Последовательностей Двойного охвата". Журнал неврологии. 28 (40): 10081–10089. doi:10.1523/jneurosci.3423-08.2008. ЧВК 2744218. PMID 18829966.
10. ^ Ван Миер Х. И., Перлмуттер Дж. С., Петерсен С. Е. (2004). "Функциональные изменения в мозговой деятельности во время усвоения и отработки последовательностей движений". Управление двигателем. 8 (4): 500–520. doi:10.1123/mcj.8.4.500. PMID 15585904.
11. ^ Уандер Дж. Д., Блейкли Т., Миллер К. Дж., Уивер К. Э., Джонсон Л. А., Олсон Дж. Д., Оджеманн Дж. Г. (2013). "Распределенная адаптация коры головного мозга во время обучения задаче интерфейса мозг-компьютер". Труды Национальной академии наук. 110 (26): 10818–10823. doi:10.1073/pnas.1221127110. PMC 3696802. PMID 23754426.
12. ^ Солсо, Роберт (февраль 2001). "Активность мозга у опытного и начинающего художника: Исследование фМРТ". Леонардо. 34 (1): 31–34. doi:10.1162/002409401300052479. S2CID 7126922.
13. ^ Дин Х., Хаган М., Песаран Б. (2012). "Только Когерентный всплеск в задней теменной коре Координирует поиск и достижение". Нейрон. 73 (4): 829–841. doi:10.1016/j.нейрон.2011.12.035. ЧВК 3315591. PMID 22365554.
14. ^ Куанг, С.; Морел, П.; Гейл, А. (2016). "Планирование движений в зрительном и физическом пространстве в задней теменной коре обезьян". Кора головного мозга. 26 (2): 731–747. doi:10.1093/cercor/bhu312. PMID 25576535.
15. ^ Берманн М., Ген Дж. Дж., Шомштейн С. (2004). "Теменная кора и внимание". Современное мнение в нейробиологии. 14 (2): 212–217. doi:10.1016/j.conb.2004.03.012. PMID 15082327. S2CID 7789667.
16. ^ Малхотра П., Култхард Э. Дж., Хусейн М. (2009). "Роль правой задней теменной коры в поддержании внимания к пространственным местоположениям с течением времени". Мозг. 132 (3): 645–660. doi:10.1093/мозг/ость350. PMC 2664449. PMID 19158107.
17. ^ Беррихилл М. Э., Олсон И. Р. (2008). "Участвует ли задняя теменная доля в восстановлении рабочей памяти? Данные, полученные от пациентов с двусторонним повреждением теменной доли". Нейропсихология. 46 (7): 1767–1774. doi:10.1016/j.нейропсихология.2008.01.009. ЧВК 2441642. PMID 18308348.
18. ^ Урожденная Д. Э., Браун Дж. У. (2012). "Диссоциируемые Лобно-полосатые и Лобно-теменные сети, участвующие в обновлении иерархических контекстов в рабочей памяти". Кора головного мозга. 23 (9): 2146–2158. doi:10.1093/cercor/bhs194. ЧВК 3841420. PMID 22798339.
19. ^ Wendelken C (2015). "Метаанализ: как задняя теменная кора способствует рассуждению?". Границы в области нейробиологии человека. 8: 1042. doi:10.3389/fnhum.2014.01042. ЧВК 4301007. PMID 25653604.
20. ^ Виттинг Н, Куперс РК, Свенссон П, Арендт-Нильсен Л, Йедде А, Йенсен ТС (2001). "Экспериментальная аллодиния, вызванная кистью, активирует заднюю теменную кору". Неврология. 57 (10): 1817-24. doi:10.1212/wnl.57.10.1817. PMID 11723270. S2CID 8586536.
21. ^ Десмерджет М., Рейли К. Т., Ричард Н., Сатмари А., Моттоле С., Сиригу А. (2009). "Намерение Движения После Стимуляции Теменной Коры у людей". Наука. 324 (5928): 811-813. doi:10.1126/наука.1169896. PMID 19423830. S2CID 6555881.
22. ^ Хаггард П. (2009). "Источники человеческой воли". Наука. 324 (5928): 731-733. doi:10.1126/наука.1173827. PMID 19423807. S2CID 206519896.
Внешние ссылки[править]
- Электрическая стимуляция вызывает чувство свободы воли
скрыть
| |||||||||||||||
| Лобная доля |
| ||||||||||||||
| Теменная доля |
| ||||||||||||||
| Затылочная доля |
| ||||||||||||||
| Височная доля |
| ||||||||||||||
| Межлобковые борозды/трещины |
| ||||||||||||||
| Лимбическая доля |
| ||||||||||||||
| Островковая кора головного мозга |
| ||||||||||||||
| Общая информация |
| ||||||||||||||
| Некоторые классификации являются приближениями, а некоторые области Бродмана охватывают извилины. |
https://en.wikipedia.org/wiki/Posterior_parietal_cortex