ФУНКЦИОНАЛЬНЫЕ СТРУКТУРЫНЕЙРОНА
В структуре нейрона выделяют элементы, которые отличаются между собой по анатомическому строению и по выполняемым функциональным обязанностям.
Нейрон состоит из:
· тела нервной клетки;
· дендритов;
· аксона;
· синапса;
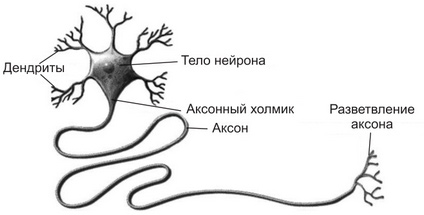
Рис. 1. Строение нейрона.
Тело (сома, перикарион) нейрона содержит ядро и органеллы, выполняет трофическую функцию, обеспечивает регенерацию аксона при повреждении, осуществляет синтез медиатора, обладает высоким порогом.
Дендриты образуют множество синапсов с другими нейронами, воспринимают информацию через рецепторы, расположенными на их мембране, обладают еще большим порогом, чем сома. На дендритах есть выросты - шипики, которые значительно увеличивают постсинаптическую поверхность дендрита. Они являются лабильными образованиями и при различных функциональных состояниях могут менять свою конфигурацию, дегенерировать и вновь появляться. В результате меняется число синапсов, меняется эффективность передачи в них нервного сигнала и т.д.
Дендриты:
• м.б. более 1мм длиной;
• занимают более 90% поверхности тела;
• проксимальные части Д. – содержат тельца Ниссля и аппарат Гольджи;
• большую часть Д. занимают микротрубочки и нейрофиламенты;
• обладают потенциал-зависимым проведением импульса (связано с Са-каналами;
Аксон берет начало от тела; начальный сегмент аксона (аксонный холмик) обладает очень низким порогом и является пусковой зоной для возникновения потенциала действия (ПД); аксон заканчивается множеством коллатералей, терминали которых образуют синапсы с другими клетками; выполняет функцию проведения ПД, таким образом осуществляя связь между нейронами и эффекторными органами.
Пресинаптическое окончание аксона содержит нейросекреторный аппарат для передачи возбуждения на другие нейроны или эффекторные клетки.
Мембрана аксона содержит специфические рецепторы к различным медиаторам или нейромодуляторам, поэтому процесс выделения медиатора пресинаптическими окончаниями может модулироваться другими нейронами.
Возникновение ПД в области аксонного холмика (возбудимость в 2 раза выше) позволяет распространяться возбуждению по аксону и телу клетки. Это важно для взаимосвязи различных частей нервной клетки и крайне важно для осуществления интегративной функции нейрона - способности суммировать влияния, поступающие к нейрону по разным синаптическим путям.
Процесс аксонного транспорта играет роль в перемещении вируса полиомиелита из ЦНС на периферию. Ретроградный транспорт, в свою очередь, играет роль в транспорте столбнячного токсина из раны в ЦНС.
Аксонный транспорт – это перемещение по аксону различного биологического материала. Аксоны являются каналами для транспорта различных веществ от тела клетки к его окончанию и обратно.
Два вида аксонного транспорта:
· Быстрый;
· Медленный.
Быстрый антероградный – это транспорт везикул, митохондрий и некоторых белковых частиц со скоростью 250-400 мм/сутки от тела клетки к терминалям аксона.
Быстрый ретроградный (от терминалей аксона к телу клетки) перемещает лизосомы, везикулы, возникающие в процессе пиноцитоза (холинэстеразы, токсинов, вирусов) со скоростью 220 мм/сутки.
Медленный транспорт обеспечивает перемещение со скоростью 1-4 мм/сутки белков и структур цитоплазмы в дистальном направлении; имеет особое значение в процессах роста и регенерации отростков нейрона.
Свойства нейрона:
· Раздражимость
· Возбудимость.
· Рефрактерность.
· Проводимость.
· Лабильность нейрона - это скорость протекания элементарных реакций, лежащих в основе его возбуждения.
Нейрон способен разряжаться с определенной частотой, которая определяется длительностью следовой гиперполяризации ПД. Чем меньше длительность следовой гиперполяризации, тем больше частота разрядов нейрона.
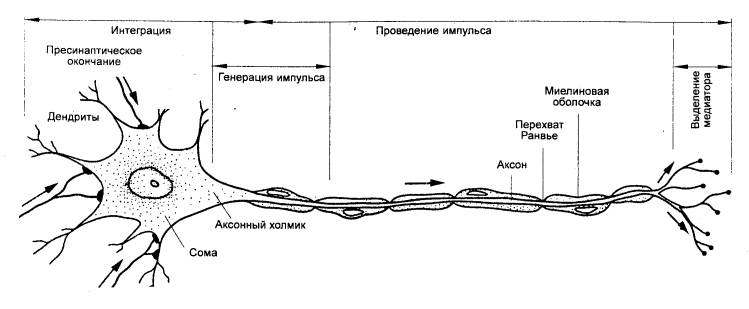
Рис. 2.Мотонейрон спинного мозга позвоночных. Функции разных его частей.
Глиальные клетки ЦНС.
Глиальные клетки – это отростчатые клетки. У глиальных клеток отсутствует аксон. Клетки глии представляют собой структуры с активным метаболизмом, которые содержат обычные органеллы, включая митохондрии, эндоплазматический ретикулум, рибосомы, лизосомы, а также отложения гликогена и жира.
Четыре основные разновидности глиальных клеток:
1. астроциты - расположены между кровеносными сосудами, телами нейронов и служат компонентом гематоэнцефалического барьера, обеспечивают опорную функцию, репаративные процессы нервных стволов, изолируют нервное волокно, участвует в метаболизме нейронов;
2. олигодендроциты – это клетки, имеющие один отросток; обеспечивают опорную, защитную функцию;
3. микроглия - самые мелкие клетки глии, являются фагоцитами мозга;
4. шванновские клетки - расположены в периферических нервах, изолируют аксон;

Рис.3.Некоторые виды глиальных клеток.
Функции глиальных клеток:
· являются опорным, защитным и трофическим аппаратом для нейронов;
· поддерживают определенную концентрацию ионов Ca и K в межклеточном пространстве;
· активно поглощают медиаторы, ограничивая время их действия.
КЛАССИФИКАЦИЯ НЕЙРОНОВ
Классификации нейронов:
I. По количеству отростков различают нейроны:
· Униполярные;
· Псевдоуниполярные;
· Биполярные;
· Мультиполярные нейроны;

Рис. 4. Классификация нейронов по количеству отростков.
II. По функции различают нейроны:
· Афферентные (чувствительные, сенсорные) нейроны, которые воспринимают стимулы из внешней и внутренней среды;
· Вставочные (ассоциативные, интернейроны) нейроны, которые связывают нейроны друг с другом;
· Эфферентные (двигательные) нейроны, которые передают возбуждение к исполнительным органам (эффекторным органам).
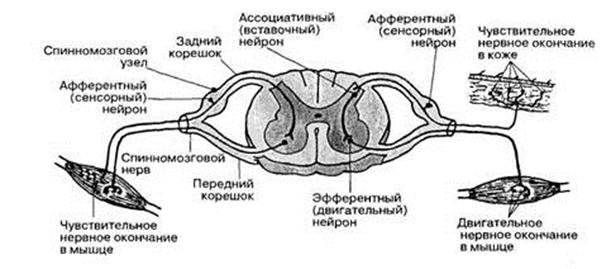
Рис. 5. Схема Рефлекса.
III. По физиологическому действию различают нейроны:
· Возбуждающие;
· Тормозные;
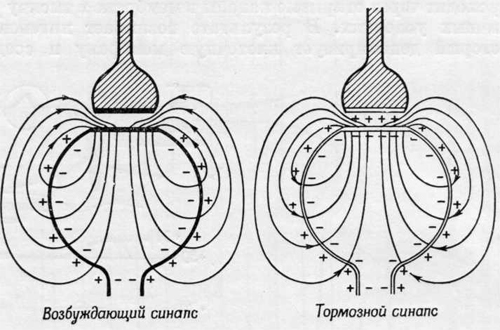
Рис. 6. Линии тока в возбуждающем (слева) и тормозном (справа) синапсах.
IV. По выделяемому медиатору различают нейроны:
· Адренергические;
· Холинергические;
· Серотонинергические и др.
Типы нейронов:
A) пирамидальная форма сомы (Н. коры головного мозга);
B) округлая сома, выраженная сеть Д. на одном конце и аксон на другом - клетки Пуркинье (Н. коры мозжечка);
C) симпатические постганглионарные Н.
D) мотонейроны спинного мозга (C и D Н. – мультиполярные Н. с радиально расположенными Д.);
E) чувствительные Н. дорзальных ганглиев – не имеют Д., аксон делится на центральную и периферическую ветви (псевдоуниполярные нейроны);

Рис. 7. Типы нейронов: а - псевдоуниполярный нейрон; б - биполярный нейрон; в - мотонейрон спинного мозга; г - пирамидный нейрон коры больших полушарий; д - клетка Пуркинье мозжечка; 1 - дендрит; 2 - тело нейрона; 3 —аксон; 4 - коллатераль аксона
Функции нейронов:
I. Неспецифические функции нейронов:
· обмен веществ;
· энергетическая функция;
· поддержания своей структуры;
· способность к адаптации;
· регулирующее влияние на соседние клетки;
II. Специфичные функции нейронов:
· получение информации от других клеток через синапсы;
· переработка и хранение информации;
· передача информации другим клеткам через синапсы (согласно классическим представлениям этот процесс осуществляется по аксону, хотя имеются данные, что выходные единицы встречаются и на дендритах).
МЕХАНИЗМ ВОЗБУЖДЕНИЯ НЕЙРОНОВ
Механизм передачи возбуждения в химических синапсах ЦНС в общих чертах таков же, как и в нервно-мышечном синапсе, но имеется ряд отличительных особенностей.

Рис. 8. Изменение возбудимости мембраны нервного волокна в различные фазы потенциала действия
1. В возникновении ПД в нейронах в отличие от нервных и мышечных волокон (скелетной мышцы) принимают участие ионы Са2+, ток которых в клетку более медленный, чем ток Na+. В частности, в дендритах клеток Пуркинье мозжечка выявлены не только быстрые натриевые потенциалы, но и медленные кальциевые, вход Са+ в пресинаптических окончаниях обеспечивает выброс медиатора, вход Са+ в дендриты нейрона примерно равен входу Na+ в тело нейрона при возбуждении. В телах некоторых нервных клеток ПД создается преимущественно за счет Са2+, а в аксоне - главным образом за счет Na+.
2. Для возбуждения нейрона (возникновения ПД) необходимы потоки афферентных импульсов и их взаимодействие. Один пришедший к нейрону импульс вызывает небольшой возбуждающий постсинаптический потенциал (ВПСП) - всего 0,05 мВ (миниатюрный ВПСП). Одновременно могут возникать не только возбуждающие, но и тормозные потенциалы. Один пузырек (квант медиатора) содержит 110 тыс. молекул медиатора. Один ПД, пришедший в пресинаптическое окончание, обеспечивает выделение 200-300 квантов медиатора. Пороговый потенциал нейрона 5-10 мВ, следовательно для возбуждения нейрона требуется некоторое множество импульсов. Выброс медиатора из нервного окончания обеспечивает входящий в деполяризованную терминаль ток Са2+, причем количество медиатора прямо пропорционально входу ионов Са2+. Действует Са2+ с помощью белка - кальмодулина, при этом четыре иона Са+ обеспечивают выброс одного кванта медиатора. При поступлении импульсов к нейрону-мишени в результате суммации ВПСП различных входов возникает деполяризация генераторного пункта в нейроне (аксонныйхолмик) которая, достигнув критической величины, обеспечивает возникновение ПД нейрона-мишени. ВПСП возникает вследствие суммарного тока в клетку и из клетки различных ионов согласно электрохимическому градиенту через ионные каналы, функциональная активность которых определяется присутствием медиатора. Поступивший в пресинаптическое окончание Са2+ удаляется за его пределы с помощью Са-насоса. Прекращение действия выделившегося в синаптическую щель медиатора осуществляется частично посредством обратного захвата его пресинаптическим окончанием, частично - с помощью разрушения специальными ферментами. Норадреналин расщепляется моноаминоксидазой и катехолметилтрансферазой, ацетилхолин гидролизуется ацетилхолинтрансферазой, имеющейся в синаптической щели и встроенной в постсинаптическую мембрану. Прекращение действия избытков медиатора на постсинаптическую мембрану предотвращает десенситизацию - снижение чувствительности постсинаптической мембраны к действующему медиатору. Медиаторы, ферменты, белки, митохондрии транспортируются в пресинаптические окончания из тела клетки по аксону с помощью микротрубочек и микрофиламентов, тянущихся по всей длине аксона. Для этого транспорта необходимы Са2+ и энергия (АТФ непрерывно ресинтезируется в аксоне). Из синапса ретроградно транспортируются по аксону в тело клетки вещества, регулирующие в ней синтез белка.
3. Место возникновения генераторных ВПСП, вызывающих ПД нейрона. Подавляющее большинство нейрональных синапсов находится на дендритах нейронов, в частности в коре большого мозга, согласно расчетам, 98 % и только 2 % - на телах нейронов. Площадь мембраны тела нейронов на 40 % занята синапсами, дендритов - на 75 %. Отношение числа синапсов к нейронам в коре большого мозга составляет 40 000:1. Наиболее эффективно вызывают возбуждение нейрона синаптические контакты, расположенные на теле нейрона. Это связано с тем, что постсинаптические мембраны этих синапсов располагаются в непосредственной близости от места первичного возникновения ПД, располагающегося в аксонном холмике. Близость соматических синапсов к аксонному холмику обеспечивает участие их ВПСП в механизмах генерации ПД. В связи с этим некоторые авторы предлагают называть их генераторными синапсами.
4. Место возникновения ПД. Генераторный пункт нейрона - аксонный холмик. Синапсы на нем отсутствуют. Отличительной особенностью мембраны аксонного холмика является высокая ее возбудимость, в 3-4 раза превосходящая возбудимость сомадендритной мембраны нейрона, что объясняется более высокой концентрацией Na-каналов на аксонном холмике. ВПСП электротонически достигают аксонного холмика, обеспечивая здесь уменьшение мембранного потенциала до критического уровня. В этот момент возникает ПД. Возникший в аксонном холмике ПД, с одной стороны, ортодромно переходит на аксон, с другой - антидромно на тело нейрона (рис. 3). Поскольку постоянная длины мембраны нейрона составляет 1-2 мм (расстояние, на котором ВПСП уменьшается на 37 %), а диаметр тела нейрона в несколько десятков раз меньше, то величина ВПСП, достигающая аксонного холмика, достаточна для возникновения ПД на нем.

Рис. 9. Регистрация (а) и временное течение ПД и ВПСП (б) в мотонейроне спинного мозга позвоночного при одиночной стимуляции заднекорешковых волокон: 1 - начальный сегмент аксона; 2 – тело нейрона; 3 - синапс; 4 – регистрирующий электрод.
При возбуждении нейронов потребление О2 возрастает в 2 раза, уменьшается количество нуклеиновых кислот в цитоплазме (иногда в 5 раз). Источником энергии является в основном глюкоза крови, собственные небольшие запасы гликогена достаточны лишь на 3-5 мин работы нейрона.
5. Роль дендритов в возникновении возбуждения до сих пор дискутируется. Дендритные синапсы удалены на значительное расстояние от генераторного пункта нейрона. По этой причине их ВПСП не могут вызвать там должной деполяризации и обеспечить генерацию ПД. Считают, что синаптический аппарат дендритов проявляет себя при одновременном поступлении возбуждения к значительному числу дендритных синапсов, при этом суммарный дендритный ВПСП, изменяя мембранный потенциал генераторного пункта на подпороговом уровне электротонически, вызывает лишь модуляцию его возбудимости, делая возбудимость большей или меньшей в зависимости от временных и амплитудных характеристик колебаний мембранного потенциала генераторного пункта относительно величины критического уровня деполяризации. Данное обстоятельство, как выяснилось, может отразиться на выраженности ответной реакции нейрона при поступлении к нему в этот момент возбуждений через синапсы тела нейрона. В связи с этим дендритные синапсы получили название модуляторных синапсов.