ЦЕНТРАЛЬНОЕ ТОРМОЖЕНИЕ
ПО И.М.СЕЧЕНОВУ [[1]]
История развития учения о тормозных процессах в центральной нервной системе начинается с открытия И.М.Сеченовым эффекта центрального торможения [V.G.4].
При нанесении раздражения на промежуточный мозг (зрительные бугры) лягушки наступает резкое торможение спинномозговых рефлексов. Этот факт, установленный И.М.Сеченовым в 1862 г., был первым доказательством наличия центрального торможения. В основе этого эффекта лежит, по-видимому, нисходящее тормозящее влияние ретикулярной формации, осуществляемое по механизму пресинаптического торможения. Мотонейроны сгибателей лапки не возбуждаются центростремительными импульсами от кожных рецепторов, но отвечают на импульсы, поступающие по другим афферентным путям.
У крупной лягушки вскрывают полость черепа и глазным скальпелем производят разрез по нижнему краю больших полушарий, отделяя их от промежуточного мозга

Рис. Головной мозг лягушки:
1 — большие полушария мозга,
2 — зрительные бугры,
3 — мозжечок,
4 — ромбовидная ямка

Подвесить лягушку за нижнюю челюсть на крючке штатива и дважды (с промежутком в 2 мин) определим время рефлекса, опуская лапку в стаканчик с 0,3%-ным раствором серной кислоты.

Рис. Ватными тампончиками и кусочками фильтровальной бумаги тщательно обсушить поверхность разреза мозга и наложить кристалл поваренной соли на зрительные бугры и определить время рефлекса. Оно значительно замедляется.
Удалить кристаллик соли, обмыть поверхность мозга раствором Рингера и обсушить ее. Через 3 и 6 мин снова определить время рефлекса.
Действие стрихнина на центральную нервную систему
Алкалоид стрихнин обладает специфическим влиянием на нервные центры спинного мозга. Он блокирует расположенные на нервных клетках тормозные синапсы, в результате чего устраняется постсинаптическое торможение. Возбудимость нервных центров при этом резко повышается — координация их деятельности нарушается. Даже очень слабые раздражители вызывают сильнейшее общее возбуждение, которое ведет к судорожному сокращению мышц. Копировать стрихнинные судороги можно введением наркотиков, в частности барбамила.
Двум лягушкам в спинной лимфатический мешок ввести по 0,5—1,0 мл 0,0,2 %-ного раствора стрихнина: второй лягушке, кроме того, ввести под кожу барбамил в дозе 7 мг на 100 г массы в виде 5 %-ного раствора. Усадить лягушек на стеклянные пластинки и закрыть большими воронками. Через каждые 1,5 ‑ 2 мин проверять их возбудимость, постукивая пинцетом по краю стекла. Первая лягушка сначала не реагирует на раздражения, затем проявляет повышенную двигательную активность, рефлекторную возбудимость, напряженность позы. Через несколько минут наступает судорожный период.
При малейшем раздражении лягушка впадает в тетанус: все мышцы тела напрягаются, задние конечности вытягиваются

Через некоторое время мышцы расслабляются, но при повторном раздражении приступ судорог повторяется с новой силой. Если поместить лягушку в стеклянную полулитровую банку, то при раздражении она вскакивает на задние конечности, оставаясь некоторое время в вертикальном положении (сокращаются и сгибатели и разгибатели, но тонус последних преобладает). У второй лягушки тонус мышц постепенно ослабляется, рефлекторная возбудимость теряется, судороги не появляются.
Декапитировать первую лягушку сначала на уровне заднего края глазниц, затем на уровне ушных капсул. Убедиться, что судороги сохраняются и исчезают только после разрушения спинного мозга.
Вначале предположение о существовании специфических тормозных нейронов, обладающих способностью оказывать тормозные влияния на другие нейроны, с которыми имеются синаптические контакты, диктовалось логической необходимостью для объяснения сложных форм координационной деятельности центральных нервных образований. Впоследствии это предположение нашло прямое экспериментальное подтверждение (Экклс, Реншоу), когда было показано существование специальных вставочных нейронов, имеющих синаптические контакты с двигательными нейронами. Активация этих вставочных нейронов закономерно приводила к торможению двигательных нейронов.
В зависимости от нейронного механизма, способа вызывания тормозного процесса в ЦНС различают следующие виды [V.G.5] торможения:
1. постсинаптическое,
2. пресинаптическое,
3. пессимальное.
Принято также различать первичное и вторичное торможение.
Первичное торможение реализуется через специфические тормозные структуры (клетки) и развивается первично без предварительного распространяющегося [V.G.6] возбуждения [++750[V.G.7] +]. К первичному торможению относятся постсинаптическое и пресинаптическое торможение.
Вторичное торможение реализуется без специфических тормозных структур (клеток) и развивается вторично после предварительного возбуждения [++750[V.G.8] +]. Т.е. торможение после возбуждения. Ко вторичному торможению относится пессимальное торможение (торможение Введенского).
Возникает вопрос: а пресинаптическое торможение разве не вторичное, ведь при этом происходит деполяризация постсинаптической мембраны аксоаксонального синапса? А ведь деполяризация – это ВПСП, а значит и возбуждение? Ответ: Нет, не вторичное: во‑первых нет распространяющегося возбуждения, во‑вторых, есть структуры специализирующиеся на торможении.
ПОСТСИНАПТИЧЕСКОЕ
ТОРМОЖЕНИЕ
Постсинаптическое торможение — основной вид торможения, развивающийся в постсинаптической мембране аксо-соматических и аксо-дендритических синапсов под влиянием активации тормозных нейронов, в концевых разветвлениях аксонных отростков.

Рис. Структуры, обеспечивающие постсинаптическое торможение.
1 — тормозный нейрон; 2 — мотонейрон.
Тормозной эффект таких нейронов обусловливается характером открываемых на постсинаптической мембране каналов (калиевых и хлорных).
Нельзя говорить: «Тормозной эффект [тормозных] нейронов обусловливается специфической природой медиатора — химического переносчика сигнала с одной клетки на другую.
При торможении в постсинаптической мембране наблюдается гиперполяризация в виде тормозных постсинаптических потенциалов (ТПСП).
Пространственно-временная суммация ТПСП повышает уровень мембранного потенциала (гиперполяризация), приводит к урежению или полному прекращению генерации распространяющихся ПД.
Наиболее распространенным тормозным медиатором является гамма-аминомасляная кислота (ГАМК).
ПРЕСИНАПТИЧЕСКОЕ
ТОРМОЖЕНИЕ [ [2] ]
Первые сведения о наличии пресинаптического торможения в ЦНС были получены в 1933 г. Г.Гассером и Г.Грэхемом, а в 1938 — Д.Барроном и Б. Мэттьюсом.[3]
Но только в 1957 г. в лаборатории Дж.Экклса[4] было показано, что амплитуда внутриклеточно регистрируемого ВПСП может уменьшиться, но при этом не регистрируется ТПСП. Авторы назвали этот тип торможения «пресинаптическим торможением» и предположили, что оно локализуется в пресинаптических волокнах.
На рис. 303220545 видно, что …
Пресинаптическое торможение развертывается в аксо‑аксональных синапсах, блокируя распространение возбуждения по аксону.
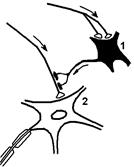
Рис. Структуры, обеспечивающие пресинаптическое торможение.
1 — тормозный нейрон; 2 — мотонейрон.
В основе его лежит процесс деполяризации постсинаптической мембраны тормозного аксо-аксонального синапса, которая именуется деполяризацией первичных афферентов (ДПА).[5] Поистине «торможение возбуждением».[6]
Ионные механизмы пресинаптического торможения еще окончательно не выяснены. [7]
Полагают, что в основе пресинаптического торможения лежит повышение проницаемости мембраны для Na+. [8]

Правда, в развитии пресинаптического торможения нельзя исключить и роли Са2++ или С1—. [9]
В отношении хлора нужно сказать, что для деполяризации мембраны этот ион должен быть выведен из клетки, а указанный процесс происходит против градиента концентрации. [10]

Значит, для его осуществления требуется «включение» соответствующего (хлорного) насоса и АТФ. [11]
ГАМКB рецепторы изучены менее детально. Расположены они на пост- и пресинаптической мембранах. Имеются данные, что посредством G‑белка ГАМКB рецепторы связаны с аденилатциклазой. Стимуляция этого подтипа рецепторов приводит к повышению содержания цАМФ, что уменьшает проницаемость ионных каналов для Са2++.

Как деполяризация (ДПА) тормозит?
Во-первых вспомните катодическую депрессию.

В связи с тем, что мембрана, где развивается пресинаптическое торможение, является частью мембраны аксона возбуждающего нейрона, возникшая волна ДПА может распространяться антидромно, навстречу нервному импульсу (ПД), идущему от сомы нейрона. [12]

Но в результате происходит не суммация, а снижение [НВГ -???] амплитуды этого ПД или полная блокада его проведения. [13]
В основе этого механизма может лежать снижение возбудимости мембраны вслед за деполяризацией — состояние рефрактерности, вызванное ДПА. И теперь более низкий потенциал действия, подходя к синапсу, вызовет высвобождение меньшего количества медиатора. [14]
Деполяризация первичных афферентов ДПА достигает своего максимума через 15-20 мс после начала и проходит через 100-150 мс.
Пресинаптическое торможение часто выявляется в структурах мозгового ствола, в спинном мозге (рис.).
ПЕССИМАЛЬНОЕ ТОРМОЖЕНИЕ
Пессимальное торможение представляет собой вид торможения центральных нейронов.
Оно наступает при высокой частоте раздражения. В первый момент возникает высокая частота ответного возбуждения. Через некоторое время стимулируемый центральный нейрон, работая в таком режиме, переходит в состояние торможения. По типу катодической депрессии.[15]

Типы постсинаптического торможения:
1. Возвратное
2. Реципрокное
3. Латеральное
ВОЗВРАТНОЕ ТОРМОЖЕНИЕ
Возвратным торможением называется угнетение (подавление) активности нейрона, вызываемое возвратной коллатералью аксона нервной клетки.
Синоним — антидромное торможение ++414[V.G.9] +. т.е. распространяющийся в направлении противоположном нормальному.
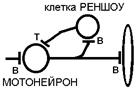
Так, мотонейрон переднего рога спинного мозга прежде чем покинуть спинной мозг дает боковую (возвратную) ветвь, которая возвращается назад и заканчивается на тормозных нейронах (клетки Реншоу). Аксон последней заканчивается на мотонейронах, оказывая на них тормозное действие.
РЕЦИПРОКНОЕ ТОРМОЖЕНИЕ
[V.G.10]


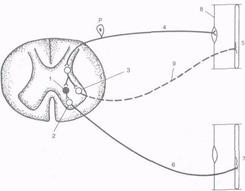
Рис. 4.3. Реципрокная иннервация мышц-антагонистов. 1 — тормозная клетка Реншоу; 2 — мотонейрон мышцы-сгибателя; 3 — мотонейрон мышцы-разгибателя; 4 — афферентное волокно от мышцы разгибателя; 5 — мышца-разгибатель; 6 — двигательное (эфферентное) волокно мышцы-сгибателя; 7 — мышца-сгибатель; 8 — мышечный рецептор; 9 — эфферентное нервное волокно мышцы-разгибателя.
ЛАТЕРАЛЬНОЕ ТОРМОЖЕНИЕ

«КОНФЛИКТ» ВОЗБУЖДЕНИЙ
Торможение спинномозговых рефлексов можно наблюдать при одновременном раздражении рецептивных полей двух рефлексов («конфликт» возбуждений). Приведём несколько примеров.
Если спинальную лягушку подвесить за нижнюю челюсть на крючке штатива. После исчезновения шоковых явлений определить время двигательного рефлекса, например, погрузив лапку в стаканчик с 0,5 %-ным раствором серной кислоты. Измерить время рефлекса. Надеть на переднюю лапку той же стороны пружинный зажим Мора и снова измерить время рефлекса. Оно резко возрастает. Снять зажим и несколько раз, с интервалом в 2—3 мин, повторить определение времени рефлекса. Оно постепенно уменьшается, возвращаясь к норме.
Если у таламического самца лягушки слегка надавливать пальцами на боковые поверхности туловища, можно вызвать квакательный рефлекс. Наложим зажим на одну из передних конечностей и повторить раздражение. Квакательный рефлекс тормозится.
Ту же лягушку положим на спину и установим, насколько быстро она переворачивается. Наденем на брюшко резиновое кольцо и повторим опыт. Рефлекс перевертывания затормаживается. Снимем кольцо — рефлекс восстанавливается.
Сделаем той же лягушке небольшой разрез брюшной полости, найдём желудок и подведём под него лигатуру, концы которой выведем наружу. Подвесим лягушку за нижнюю челюсть к штативу и определим время рефлекса, раздражая лапку 0,1 или 0,3 %-ным раствором серной кислоты. Обмоем лапку. Слегка подтянем желудок и перевяжем его лигатурой. Снова измерим время рефлекса. Оно обычно удлиняется.
СОКРАЩЕНИЯ
ДПА — деполяризация первичных афферентов (при пресинаптическом торможении). [16]
[1] [++414+ С.380]
[2] Посмотрите ещё: [2] ++743+ С.90-91 (+++); --147- С.45; ++744+ С.53-55 примерно; ++484+ С. 291-292; ++634+ С.82; ++685+ С.577-578 (+-); ++66+ С.238 плохо; ++511+ С.105-106 (плохо);
не нашёл –140-
[3] --95- С.70-72 Эти исследователи обнаружили, что при раздражении заднего корешка спинного мозга на его дорсальной поверхности регистрируется медленный позитивный потенциал, длительность которого приблизительно соответствует развитию торможения сгибательных рефлексов, вызываемых раздражением соседнего заднего корешка.
[4] К.Фрэнком и М.Фуортесом
[5] ++743+ С.90-91
[6] НВГ
[7] ++743+ С.90-91
[8] ++743+ С.90-91
[9] ++743+ С.90-91
[10] ++743+ С.90-91
[11] ++743+ С.90-91
[12] ++743+ С.90-91
[13] ++743+ С.90-91
[14] ++743+ С.90-91
[15] НВГ
[16] ++743+ С.90-91
[V.G.1] 4.1.3.
[V.G.2]++512+С.376 Алексей алексеевич Ухтомский (1875-1942) 67 лет
[V.G.3]С.115
[V.G.4](химическое раздражение зрительных бугров тормозит простые спинномозговые безусловные реакции)
[V.G.5]несколько
[V.G.6]сам дополнил, чтобы не путали с пресинаптическим торможением
[V.G.7]С.182
[V.G.8]С.182
[V.G.9]С.
[V.G.10]С.116