Морфофункциональный анализ стелющейся колонии у гидроидов на примере cordylophora inkermanica marfenin (athecata, clavidae)
Гидроид приспособлен к обитанию на.“мозаичном” субстрате (зостера, мидии) и специализирован на питании зоопланктоном. Примитивные побеги состоят всего из двух — четырех междоузлии. Их столоновидные ветви легко прилипают к субстрату, что позволяет колонии переходить с одного субстрата на другой. Длинные ветви легко отрываются от колонии. Предполагается, что первичной функцией побегов в филогенезе гидроидов было вегетативное размножение. В дальнейшем произошла смена функций — побег стал опорой для охвата колонией прилежащих ко дну слоев воды.
В морфологическом ряду усложнения организации колонии у гидроидов стелющиеся колонии занимают нижнюю ступеньку (Наумов, 1960). Стелющимися они называются потому, что гидранты в них расположены либо непосредственно на гидроризе, либо на примитивных коротеньких побегах.
Отличаясь отсутствием сложно устроенных побегов, стелющиеся колонии интересны прежде всего в двух отношениях. Во-первых, неясна особенность экологической ниши, которую занимают стелющиеся колонии, и, во-вторых, неясны первичные причины появления побегов у гидроидов. С возникновением побегов связано основное направление в эволюции гидроидов—развитые побеги позволяют им приподняться над субстратом и облавливать большее пространство вокруг себя. Однако эта экологическая эффективность побегов становится ясной лишь по мере усложнения их организации и вовсе не очевидна на этапе их возникновения, так как с маленьким побегом сопоставим крупный гидрант, что представляет собой другую линию эволюции гидроидов.
Мы вправе предполагать смену функций в процессе эволюции побегов. Изучение морфологии, роста, пищевого поведения и физиологической интеграции колониального гидроида Cordylophora inkermanica Marfenin, обнаруженного недавно в Черном море (Марфенин, 1983),.дает возможность разобраться в этом процессе, хотя бы применительно к частному случаю. Этому и посвящена настоящая статья.
Материал и метод
Cordylophora inkermanica собрана в мае 1981 г. в Севастопольской бухте вблизи Инкермана у поверхности воды с Mytilus galloprovincialis Lam. Как впоследствии выяснилось, материал был представлен исключительно женскими колониями.
Исследования биологии вида проводили частично сразу после сбора в Институте биологии южных морей (г. Севастополь), а в основном в Москве с сентября 1981 по май 1982 г.
На первом этапе в Севастополе использовали свежую черноморскую воду, которую меняли ежедневно. Температура воды была в пределах 20—25° С. Гидроиды содержали в чашках Петри без дополнительной аэрации. Кормом служили свежевылупившнеся науплиусы Artemia.
Затем гидроиды были перевезены на Беломорскую биостанцию МГУ, где их содержали в лабораторных условиях на чашках Петри в аквариумах с беломорской водой (соленость около 26%о), смену которой осуществляли дважды в неделю.
С сентября 1981 г. исследования гидроидов были продолжены в Москве. Колонии по-прежнему содержали в беломорской воде, но теперь уже только в чашках Петри, а не в аквариумах. Дополнительной аэрации и, следовательно, активного перемещения воды не было. Воду меняли один раз в неделю. Температура колебалась от 18 до 20° С. Кормление колоний в этот период было индивидуальным, т. е. из пипетки науплиусами Artemia один или два раза в сутки в зависимости от характера экспериментов.
В мае 1982 г. в Севастополе был собран дополнительный материал. На сей раз в сборах были и мужские колонии, что позволило летом 1982 г. при работе на Беломорской биостанции МГУ изучить процесс полового размножения и оседания планул при таких же условиях культивирования, как и летом 1981 г.
Для изучения роста колоний использовали методику картирования (Марфенин, 1980). Размеры и возрастные стадии гидрантов оценивали по числу сформированных щупалец.
Морфология колонии
Колонии стелющиеся (рис. 1, Б). Гидранты на ножках сидят либо непосредственно на гидроризе, либо на неправильно разветвленных небольших побегах. Гидрориза и стволики побегов покрыты перисарком. Ножки гидрантов и сами гидранты свободны от перисарка (рис. 2, 7). Ценосарк и гидранты не пигментированы, т. е. колония фактически прозрачна. Однако энтодерма может быть временно окрашена в зависимости от цвета пищи.
Гидрориза колонии нитевидная. Столоны (ветви гидроризы) в сечении округлые. Ценосарк занимает лишь часть пространства внутри перисарка и, за исключением верхушек роста, обычно в 2 раза тоньше трубки перисарка (рис. 2, 4, 5), диаметр которой 0,1 мм. Растут столоны прямолинейно. При встрече друг с другом они, как правило, анастамозов не образуют. Ветвление гидроризы происходит в междоузлиях между гидрантами, а не в основании ножек гидрантов. Боковые столоны могут возникать практически в любой части колонии. Наблюдения показывают, что это происходит обычно вблизи гидрантов, получивших много пищи. Расстояния между гидрантами (т. е. длина междоузлии гидроризы) обычно от 2,2 до 3,3 мм, хотя разброс значений в выборке из 126 измерений был от 1,9 до 6,4 мм. Эффект возрастания длины междоузлии по мере роста столонов не обнаружен. Растущие верхушки столонов, как и у других гидроидов, на конце слегка расширены (рис. 2, 3).
Гидранты формируются по мере роста столонов на их дистальных концах (рис. 1, Б, 2). Возникновение гидрантов на старых участках гидроризы не происходит. Зачаток гидранта (почка) образуется сразу за верхушкой роста столона, примерно в 0,2 мм от ее апикального конца (рис. 1, Б, 1). Почка растет и примерно через сутки (при 20° С) превращается в маленького гидранта с высотой тела около 0,2 мм с пятью-шестью короткими (0,15 мм) щупальцами. Ножка вначале слабо различима. Еще через сутки гидрант вырастает до 0,4 мм. Щупалец становится больше (8—10), а их длина достигает 0,6—0,8 мм. Появляется короткая ножка (0,4—0,5 мм), которая на самом деле лишь в дистальной части (0,2 мм) является ножкой, а в проксимальной по существу представляет собой стволик побега (рис. 1, 5, 8). В дальнейшем тело гидранта вырастает до 0,7—0,8 мм, щупальца до 2,2 мм, а их число постепенно возрастает до 14, а иногда и до 20 (рис. 2, 1).
Спустя некоторое время гидрант рассасывается, т. е. происходит его закономерная дедифференциация, хорошо известная для Campanulariidae (Crowell, 1953), с укорочением щупалец, смыканием гипостома и уменьшением длины тела полипа вплоть до полного его исчезновения (рис. 1, 5, 4). После более или менее продолжительной диапаузы на дистальном конце оставшегося стволика появляется почка, формируется новый гидрант, а в процессе его роста вырастает и сам стволик побега (рис. 1, 5, 5). Гидрант второй генерации по форме и размерам не отличается от гидранта первой генерации.
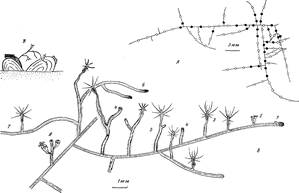
Рис. 1.А—пример карты колонии средней величины. Линии — гидрориза, белые кружки—функционирующие гидранты, черные—рассосавшиеся гидранты; Б—фрагмент колонии, иллюстрирующий жизненный цикл гидранта. 1 — верхушка столона с зачатком будущего гидранта, 2—формирующийся гидрант, 3—сформированные гидранты на ножках разной длины, соответствующих возрасту гидранта, 4—стволы побегов с верхушками на месте рассосавшихся гидрантов, 5 — побеги с гидрантами второй генерации и зачатками новых гидрантов, 6 — побег со столоповидными ветвями, 7 — ветвь, прикрепившаяся к субстрату и ставшая столоном, 8 — побег с двумя спороса-ками на разных стадиях развития (гидрант рассосался); В—предполагаемое значение неправильного ветвления с образованием столоповидных побегов: “побег — мост” позволяет осваивать новые субстраты

Рис. 2. Строение гидранта, фрагмента гидроризы и верхушки столона. /—взрослый гидрант, переваривающий науплиуса Artemia, 2 — начальная стадия образования очередного гидранта — почка на ножке гидранта, 3 — верхушка столона, 4 — ценосарк столона, 5—перисарк столона. Черными точками везде показаны частицы пищи в полости колонии
Жизненный цикл гидрантов непостоянен по продолжительности (рис. 3). В 26 подробно исследованных случаях при 20° С он варьировал от 1,5 до 10,5 сут, но преимущественно был в пределах 2,5—5 сут.
Диапаузы также неодинаковы — зарегистрированы случаи их продолжительности (при нормальном питании) от 2 до 5,5 сут, но очевидно, что диапауза может быть и дольше.
Тело гидранта веретеновидной формы с коническим гипостомом, нитевидные щупальца расположены многоярусным венчиком (рис. 2, 1) и по своей морфологии похожи на таковые семейства Campanulariidae. Стрекательные клетки, как и у многих других гидроидов, сгруппированы в батареи, расположенные на щупальце спирально или периодическими венчиками, что позволяет сокращаться щупальцам в 2—3 раза.
При хорошем питании на ножке гидранта может образоваться почка (рис. 2, 2), которая будет сначала расти (рис. 1, Б, 5), а затем на ее дистальном конце сформируется гидрант. Таким образом возникают неправильно разветвленные побеги.
Побеги чаще всего состоят из одного-двух междоузлий, в чем проявляется их примитивность. Благодаря зоне роста, расположенной под гидрантом, его ножка удлиняется не только до момента сформирования гидранта из верхушки, но и после. Второе междоузлие начинается с появления боковой почки на первом. Оба междоузлия могут продолжать одинаково хорошо расти, и на каждом из них возможно появление новых боковых почек. Следовательно, по этим признакам побег должен быть отнесен к моноподиальным с зооидными зонами роста и почкования (Наумов, 1960).
Однако с каждым новым ветвлением боковая ветвь растет все дольше, прежде чем формирует гидрант на своей верхушке. Поэтому уже после третьего и тем более четвертого ветвления вместо боковой ветви образуется фактически висячий столон. Его верхушка легко прилипает к любому твердому субстрату, после чего растет, формируя настоящий столон. Вторая примитивная черта побегов в том, что часто после очередного рассасывания гидрант не возобновляется и, следовательно, эта ветвь прбега перестает расти. В результате дочерняя ветвь может обогнать материнскую. Получается примитивно моноподиальный побег с кривыми ветвями разной длины, расположенными слабоупорядочепно (рис. 4). Получить колонии с такими побегами удавалось при культивировании гидроида в течение нескольких месяцев в неподвижной воде. Несмотря на малое число междоузлий, побеги оказываются довольно протяженными—до 1—2 см. Они не способны вертикально возвышаться над субстратом.
Часто в проксималыюй части такого побега, переросшего в висячий столон, ценосарк истончается, а иногда и рвется. Физиологическая взаимосвязь между побегом и остальной колонией нарушается, однако он остается жизнеспособным благодаря тому, что на нем есть несколько функционирующих гидрантов. Длинные плетевидные побеги легко отрываются от колонии, чему, вероятно, способствует утончение цепосарка в их основании. Оторванный побег сохраняет способность прикрепляться к субстрату своей верхушкой. Поэтому образование и отрыв побегов можно рассматривать как один из способов вегетативного размножения.
Споросаки образуются на стволиках побегов обычно по два-три, но их бывает и до шести одновременно. Сформированные мужские споросаки яйцевидной формы (рис. 5, 1) и заполнены однородным содержимым. Женские споросаки в начале своего развития почти сферические (рис. 5, 2), а затем становятся грушевидными. В дистальной расширенной части споросака видны яйцеклетки (рис. 5, 2), расположенные вокруг спадикса одним слоем. Формирующиеся планулы сначала шаровидные, около 0,12 мм в диаметре, и через некоторое время оказываются вывернутыми над апикальным концом споросака (рис. 5, 3), а еще спустя некоторое время они вытягиваются, становятся подвижными и уплывают (рис. 5, 4). Дальнейшее их поведение обычно: после непродолжительного периода плавания в толще воды, в течение которого они вертятся вокруг собственной оси, планулы идут на дно и начинают ползать по субстрату со скоростью до 0,75 мм/с.

Рис. 3. Вариационный ряд продолжительности жизненного цикла гидрантов в колонии. Всего рассмотрено 26 случаев. По оси ординат — число гидрантов (N} с данной продолжительностью жизненного цикла, по оси абсцисс — продолжительность жизненного цикла (сут). Объяснения в тексте

Рис. 4. Пример большого побега с неупорядоченным строением и столоновидными ветвями
Передний конец их совершает рыскающие движения. Периодически планула изгибается в виде арки, пытаясь прикрепиться к субстрату (рис. 5, 5). В случае удачи она становится вертикально и через некоторое время образует первичный гидрант (рис. 5, 6) высотой около 0,3 мм и с четырьмя щупальцами короче 0,1 мм. Весь процесс оседания планулы занимает менее суток. Первичные гидранты должны питаться простейшими, так как проглотить свежевылупившегося науплиуса Artemia они не могут, хотя способны заякорить и присосаться к нему. Через 4 сут при наличии корма первичные гидранты вырастают до 0,5—0,7 мм высотой и имеют шесть—восемь щупалец, т. е. достигают обычных размеров молодых гидрантов.
Количественные показатели структуры колонии. Количественные соотношения числа или размеров гомономных частей колонии друг с другом называются показателями структуры колонии (Марфенин, 1977). Они отражают специфику колониального строения — ограниченность разнообразия частей, из которых построена колония (например, гидранты, гонофоры, побеги, гидрориза, верхушки роста) при одновременной множественности гомономных частей.
У С. inkermanica эти показатели имеют следующие примерные значения: 1)H/L — отношение числа всех гидрантов (формирующихся,, сформированных и рассосавшихся) к длине гидроризы, выраженной в мм. Показатель дает представление о возможной наивысшей частоте расположения гидрантов в данной колонии. В связи с рассасыванием части гидрантов реальная частота расположения функционирующих в момент наблюдения гидрантов меньше и выражается показателем Hf/L, где Hf—число функционирующих гидрантов. В выборке из 22 случаев (далее выборка та же) H/L=0,33—0,48 при среднем 0,38, Hf//L=0,09—0,30 при среднем 0,21 (гидрант/мм).
B/L — отношение числа всех зон роста (верхушки роста гидроризы, побегов, формирующиеся гидранты, удлиняющиеся ножки функционирующих гидрантов) к длине гидроризы (в мм). Показатель дает представление о частоте расположения зон роста в колонии. B/L=0,16— среднее: 0,38; Bl/L—отношение числа растущих верхушек гидроризы (Bl) к длине гидроризы (в мм).

Рис. 5. Половое размножение, 1 — мужские споросаки, 2, 2'-— женский споросак, 3 — развивающиеся планупы на женских споросаках, 4—планула, 5 — прикрепляющаяся планула, 6— первичный гидрант
3) HfIB — отношение числа функционирующих гидрантов (Hf) к числу всех зон роста. Показатель косвенно дает представление о среднем количестве пищи, приходящемся на одну зону роста. В данном случае этот показатель довольно точно соответствует числу науплиусов
Artemia, полученных колонией за 12 ч в расчете на одну зону роста (см. ниже эксперимент с пропорциональным кормлением). Hf//В=0,40—0,77, среднее: 0,56.
4) HdIL—отношение числа рассосавшихся гидрантов (На) на момент наблюдения к длине гидроризы (в мм). Это показатель интенсивности рассасывания гидрантов в колонии, и он сопоставим с показателями H/L и Hf/L. В рассматриваемом случае Hд/L=0,03—0,26, среднее: 0,11
РОСТ КОЛОНИИ
Рост при пропорциональном кормлении. Изучение роста колонии при режиме пропорционального кормления, когда она получает количество пищи, пропорциональное числу функционирующих гидрантов, весьма трудоемко, так как колония, имея достаточное (или даже избыточное) количество пищи, растет очень быстро, вследствие чего ее картирование затрудняется с каждым последующим днем.
В данном случае был прослежен рост всего одной колонии в течение 11 сут, но зато наиболее подробно. Карты колонии составляли дважды в сутки: в 10 и 22 ч- Это позволило точнее (чем при обычном картировании через 4 или 2 сут) определить скорости роста столонов, а также проследить как динамику ветвления столонов и побегов, так и жизненный цикл гидрантов. Кормление осуществляли дважды в сутки из пипетки свежевылупившимися науплиусами Artemia. Каждый гидрант получал, как правило, по одному науплиусу. Следовательно, пропорциональное кормление не было в данном случае неограниченным.
За 11 сут колония выросла примерно в 6 раз. Протяженность гидроризы от начальной (40,7 мм) увеличилась до 266,5 мм, а число всех гидрантов возросло от 18 до 93. Всего за это время колония получила 436 науплиусов. Таким образом, прирост гидроризы в расчете на одного науплиуса составил 0,5 мм, а прирост числа гидрантов—0,17 гидранта/ /науплиус.
Рост колонии можно подразделить па три этапа (рис. 6). На первом этапе он идет медленно, затем ускоряется и на втором этапе (с 7 по 11 февраля) приближается к экспоненциальному. Последовавшее на третьем этапе замедление роста объясняется закономерным рассасыванием старых гидрантов и наступлением периода депрессии. Продолжавшиеся после завершения картирования наблюдения позволили установить, что после периода депрессии наступил очередной этап бурного роста.
Прирост отдельных столонов за 12 ч варьировал от 0,1 до 2,3 мм, но чаще всего он был около 1,0 мм, минимальные значения его—менее 0,5 м за 12 ч—относятся к периоду депрессии, когда скорость роста столонов уменьшается. Основные или длинные столоны растут, как правило, несколько быстрее (0,9—1,6 мм за 12 ч), чем короткие боковые (0,3—1,2 мм за 12ч).
У отсеченных частей колонии проксимальный конец столона, как правило, не растет. Рост в этой части колонии может осуществляться за счет образования боковых столонов. Тот же эффект полярности столона наблюдается и при автофрагментации гидроризы в периоды глубокой депрессии (см. ниже): в месте разрыва ценосарка столона его рост возобновляется активнее с одной стороны, для которой место разрыва является дистальным.
Депрессии роста колонии. Уменьшение числа функционирующих гидрантов обуславливает дефицит пищи, а это в свою очередь приводит к уменьшению числа зон роста. Вначале прекращается ветвление побегов и гидроризы, а затем приостанавливается образование очередных генераций гидрантов взамен рассосавшихся на старых побегах. Последнее лишь обостряет дефицит пищи, тем более что основные столоны в колонии продолжают расти.
Наступает период депрессии: число функционирующих гидрантов сокращается, начинается рассасывание дистальных частей побегов, скорость роста столонов падает. Затем некоторые столоны прекращают рост. Ценосарк гидроризы истончается повсеместно, но особенно сильно на некотором расстоянии от растущих верхушек столонов. Депрессии бывают различной степени. При глубокой депрессии возможно рассасывание всех гидрантов в колонии. В этом состоянии обычно наступает автофрагментацня гидроризы—разрывы ценосарка столонов в наиболее истонченных местах. На некотором расстоянии от верхушек столонов разрывы бывают многочисленными, так что в дистальной части столона остается цепочка коротких фрагментов, большая часть которых нежизнеспособна. Реже единичные разрывы происходят в проксимальных областях столонов, недалеко от их ответвления от материнского столона. Если в первом случае автофрагментация фактически уничтожает верхушку роста столона, то во втором изолированной оказывается полноценная часть колонии. Поэтому можно считать второй вариант автофрагментации одним из способов бесполого размножения колонии.

Рис. 6. Увеличение длины гидроризы (1), числа, функциони рующих гидрантов (2) и рассосавшихся гидрантов (3) у одной колонии во время эксперимента с пропорциональным кормлением (февраль 1982 г.). По оси абсцисс—дни, по оси ординат: A — длина гидроризы (мм) и число функционирующих гидрантов; Б—число рассосавшихся гидрантов
Выход из состояния депрессии начинается с появления молодых гидрантов на дистальных концах столонов основной колонии или образовавшихся фрагментов. По-видимому, получаемой ими пищи после значительного сокращения числа зон роста хватает для обеспечения роста оставшихся. За счет этого формируются новые гидранты, что приводит к улучшению питания колонии. Возобновляется рост некоторых из остановившихся столонов. Ценосарк колонии постепенно возвращается в нормальное состояние: утолщается и окрашивается в цвет пищи. В некоторых местах разрыва ценосарка восстанавливается его целостность. Выход из состояния депрессии идет тем быстрее, чем меньше размеры колонии. По-видимому, это связано с энергетическими затратами на поддержание существования колонии, которые тем выше, чем больше ее размеры. Поэтому изолировавшиеся во время депрессии фрагменты колонии “оживают” быстрее, чем сама материнская колония.
Дальнейшее восстановление колонии после депрессии происходит все быстрее и быстрее. При избытке пищи, который получается вследствие возрастания числа гидрантов, начинается ветвление гидроризы вблизи мест получения пищи. Это в свою очередь приводит к еще большему избытку пищи, так как за счет увеличения числа растущих столонов сразу образуется значительно больше новых гидрантов. Возобновляется рост старых побегов. У них на местах рассосавшихся ранее гидрантов образуются новые. Затем начинается ветвление побегов.
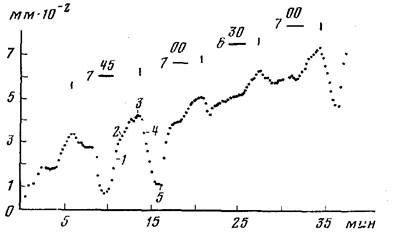
Рис. 7. Пример характерных пульсаций верхушки столона. Регистрация по апикальной части верхушки через 24 ч после кормления колонии. Температура воды 21,5° С. По оси абсцисс—время, мин; по оси ординат—длина, мм-10-2. Объяснения в тексте
Одновременно постепенно нарастает доля рассасывающихся старых гидрантов. Эти два процесса—усиление роста и рассасывания гидрантов—приходят рано или поздно в противоречие друг с другом. Наступает момент, когда функционирующих гидрантов становится меньше,чем надо для обеспечения пищей всех зон роста. С этого начинается очередная депрессия. Ежедневные наблюдения с учетом числа полученных колонией науплиусов при пропорциональном кормлении в течение двух месяцев показали, что депрессии наступали через каждые 13— 15 сут. Ни с какими внешними воздействиями связать эту периодичность не удалось.
Следует, однако, отметить, что при неограниченном пропорциональном кормлении (избыточное количество науплиусов выпускается ежедневно непосредственно в сосуд с гидроидами) периодические депрессии не наблюдались.
Ростовые пульсации верхушек столонов. Пульсации верхушек роста столонов регистрировали визуально с помощью винтового окуляр-микрометра с 15-секундной периодичностью. Форма кривой ростовых пульсаций типична для большинства исследованных гидроидов (рис. 7, 1—5). Этап удлинения верхушки состоит из двух фаз: быстрое возвращение к исходным размерам после сжатия (1), более медленный рост—удлинение (2). Затем наступает более или менее продолжительная фаза покоя на максимуме (3), после чего происходит достаточно быстрое укорочение верхушки (4). Завершает цикл короткая фаза покоя на минимуме (5).
Амплитуда пульсаций зависит от ряда факторов, в том числе от гидроплазматического давления в полости верхушки. Увеличение давления, которое происходит при сжатии других участков колонии, приводит к резкому уменьшению амплитуды, наблюдаемому в некоторые циклы. Пульсации периодичны. Зарегистрированный в нашем случае период при 21,5° С был от 6 мин 30 с до 7 мин 45 с. Специальных исследований зависимости пульсаций от температуры, солености и других факторов не проводили.
Пищевое поведение гидрантов и переваривание пищи в них
В спокойной воде гидранты обычно неподвижны. Им не свойственны ни изгибания щупалец или тела, как у Tubularia или Clava, ни периодические схлопывания всех щупалец вместе, как у Оbеlia или Dynamena.
Заякоривание науплиуса Artemia происходит с первого же раза любыми щупальцами. Только гидранты, начавшие рассасываться, хуже заякоривают добычу. Удержание жертвы и подтягивание ее к гипостому происходит одним или несколькими щупальцами. Остальные щупальца могут быть в это время несколько сжаты или даже отведены назад вдоль тела гидранта. Гипостом открывается навстречу жертве и натягивается на нее. Щупальца в заглатывании не принимают участия. Время проглатывания свежевылупившегося науплиуса взрослым голодным гидрантом обычно от 4 до 8 мин, в зависимости от положения науплиуса при заглатывании. Время проглатывания учитывали от момента заяко-ривания жертвы до смыкания гипостома после заглатывания.
Обычно гидрант может проглотить одного (редко двух) науплиуса, хотя заякорить может нескольких. Щупальца полностью возвращаются в исходное ловчее положение через 5—10 мин после проглатывания науплиуса. У старого или сытого гидранта щупальца могут более 2ч оставаться в нерабочем положении. Во время заглатывания жертвы и после этого свободные щупальца способны продолжать заякоривать новые жертвы. В этом случае после заякоривания жертва висит некоторое время обездвиженная на щупальце, а затем спадает.
Молодые гидранты, не достигшие взрослых размеров, также хорошо заякоривают науплиусов Artemia, но проглотить их не могут. В этом варианте гипостом маленького гидранта выворачивается наружу, и гидрант присасывается к науплиусу. Очевидно, таким образом гидранты также способны питаться, хотя науплиус на первый взгляд остается непереваренным. Чуть более крупные гидранты способны заглотить часть добычи и переваривать ее; остальная часть остается непереваренной и затем отбрасывается.
Переваривший только что пищу гидрант заглатывает второго науп лиуса дольше, чем первого. Старые гидранты на последних этапах своего жизненного цикла вовсе не могут проглатывать науплиусов.
Рацион гидроидов специально не изучали, но, судя по некоторым наблюдениям, он должен быть достаточно широк. Гидранты успешно заякоривают и проглатывают икру мидий. В то же время они могут справиться и с хиропомидой, размером превышающей их в несколько раз.
Переваривание науплиуса Artemia в гидранте при температуре 20— 25° С продолжается 1—1,5 ч (в старом гидранте значительно дольше). Пищеварительная перистальтика, описанная для ряда гидроидов (Марфенин, 1981), в данном случае выражена слабо. Наблюдаются пульсации центральной зоны тела гидранта с периодом 2,5—3,5 мин, которые, возможно, являются разновидностью пищеварительной перистальтики. Пульсации тела гидранта способствуют раздавливанию пищи. В самом гидранте клетки энтодермы захватывают частицы пищи, хорошо различимые под микроскопом. Тело гидранта становится из-за этого менее прозрачным, что мешает достоверно установить момент окончания первичного переваривания науплиуса. Вывод пищи из гидранта в ценосарк колонии начинается примерно через 10—15 мин с момента окончания проглатывания добычи. Видно, как частицы или их агрегации выносятся гидроплазмой при сжатии гидранта в полость ценосарка. К этому времени контуры проглоченного науплиуса размываются. При обратном токе гидроплазмы в гидрант различимые частицы в ней чаще всего отсутствуют.
Установить время наступления экскреции не удалось, несмотря на наблюдения в течение 12 ч после кормления. По-видимому, для этих гидроидов не характерно обязательное “выплевывание” гидрантами шкурок науплиусов. В чашках Петри, в которых некоторые колонии длительно содержались в неподвижной воде, также не было обнаружено пустых шкурок науплиусов. Участки дна чашки вблизи гидрантов постепенно покрывались тонким слоем мелких частиц и их агрегаций. Удалось, однако, установить, что некоторые гидранты примерно через 10 ч после кормления (при 20° С) выворачивали гипостомы энтодермой наружу. При этом из гидранта вываливалась (или в течение нескольких часов торчала) рыхлая масса мелких частиц розовато-оранжевого цвета, входных по размерам с теми, что устилали дно. По прошествии 10—12 ч зкскпетировавшие гидранты рассасывались. Многочисленные наблюдения подтвнрждают, что вокруг рассосавшихся за последние 12 ч гидрантов, получивших до того пищу, как правило, появляются розоватые поля выбросов. Не ясно пока, можно ли считать это нормальной, экскрецией, Защитное поведение и реакция испуга сводятся у гидрантов к сжатию щупалец примерно в 2—3 раза, а иногда к отведению щупалец назад вдоль тела. Быстрого изгибания теля у гидрантов не наблюдалось.