Абсолютная чувствительность глаза
Жизненный опыт убеждает, сколь чувствителен глаз человека к свету. Астрономы давно научились краешком глаза (как мы теперь понимаем, периферическим палочковым зрением) различать на ночном небе даже самые слабые звезды. Однако необходимы были конкретные знания о минимальной энергии света или числа квантов, способных создать субъективное ощущение световой вспышки. От этого прямо зависит понимание процессов преобразования светового сигнала в зрительный, т.е. понимание молекулярных механизмов фототрансдукции.
Как мы теперь знаем, в эксперименте по определению порога чувствительности зрительной системы необходимы следующие условия: предварительная темновая адаптация глаза наблюдателя; фиксация пятна света на периферии сетчатки, где находятся более чувствительные к свету палочки (сумеречное зрение); достаточно маленькое световое пятно, падающее на сетчатку глаза, порядка 10 или менее угловых минут; кратковременная (~1 мс) световая вспышка; определенная длина волны света, соответствующая максимуму спектральной чувствительности палочкового зрения (около 510 нм).
Еще в конце XIX в. вполне грамотно определил порог чувствительности глаза американский физик и астроном С.Р.Лэнгли (1834-1906). При изучении солнечной активности ему необходимо было измерять интенсивность радиации во всем диапазоне длин волн. Так он создал тепловой детектор световой энергии - болометр, чувствительность которого не зависела от длины волны света, что и было принципиально важным для Лэнгли [1]. Современное определение гласит, что болометр - это калиброванный высокочувствительный прибор с плоской спектральной характеристкой для измерения световых потоков. Усовершенствованные болометры используются по сей день. Таким образом, директор обсерватории, профессор физики и астрономии Питсбургского университета Лэнгли вошел в историю науки как изобретатель болометра, а в историю физиологии зрения как физик, экспериментально определивший порог абсолютной световой чувствительности глаза. Согласно Лэнгли [2], значение по энергии - 3·10–9 эрг, что соответствует потоку, содержащему 800 фотонов. Это всего лишь на порядок величины выше современных значений. И это можно понять, поскольку знания физиологии зрения того времени не позволило учесть в эксперименте целый ряд факторов.
Пороговые значения, полученные независимо Ю.Б.Харитоном и С.И.Вавиловым в конце 20 - начале 30-х годов, были гораздо ближе к современным. Как и Лэнгли, для решения собственных физических задач им требовалось регистрировать исключительно слабые световые вспышки. Болометры того времени их не удовлетворяли, а других точных приборов еще не было. Самым чувствительным прибором оказывался собственный глаз экспериментатора.
Опыты Ю.Б.Харитона
Харитон, физик в жизни и гуманитарий в душе, в 1926 г., совсем молодым человеком, попал в Кембридж, в Кавендишскую лабораторию к великому Э.Резерфорду. Ему была предложена классическая задача - счет a-частиц. Тогда широко использовали сцинтилляционный метод, но, как ни странно, его характеристики были недостаточно хорошо изучены. В частности, оставалась неизвестной чувствительность метода, определяемая в первую очередь порогом зрительного восприятия. Известные к тому времени работы по нахождению абсолютной световой чувствительности глаза - минимального количества квантов, вызывающего зрительное ощущение, - давали значения в интервале от 10 до 10 000.
Харитон и его коллега С.Ли разработали психофизическую установку, в которой определение порога зрения осуществлялось в условиях повторяющихся наблюдений сцинтилляций [3]. В качестве источника света использовали газонаполненную лампу, помещенную в ящик с небольшим отверстием. Для получения достаточно однородного потока отверстие освещали светом, отраженным от медной пластинки, напыленной оксидом магния. Система из двух дисков с отверстиями, вращающихся с разной скоростью, позволяла менять длительность вспышки от 0.1 до 5·10–5с. Свет, прошедший через водяной фильтр (с полосой пропускания l = 470-520 нм), наблюдали на экране с помощью зрительной трубы. Изменение светового потока, входящего в телескоп во время вспышки, достигалось диафрагмированием объектива.
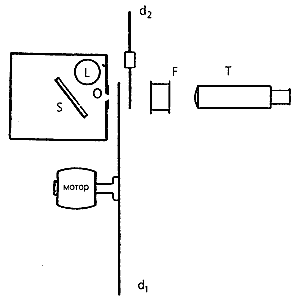
Схема установки Ю.Б.Харитона. L - газонаполненная лампа; O - небольшое отверстие в ящике; S - медная пластинка, напыленная оксидом магния; d1, d2 - диски с отверстиями, вращающиеся с разной скоростью; F - фильтр, наполненный водным раствором CuSO4 и K2Cr2O7. Подбором состава добивались минимального поглощения при l = 500 нм. Полоса пропускания фильтра (l = 470-520 нм) соответствовала цвету сцинтилляций: чувствительность глаза в этой полосе близка к максимальной. T - зрительная труба.
В опытах испытуемые довольно быстро утомлялись. Поэтому каждому из них предъявляли только по три вспышки со значительным перерывом в сериях, затем рассчитывали среднее значение по пяти повторам. Выяснилось, что тренированные по сравнению с новичками видят вспышки меньшей интенсивности. Авторы объясняли этот факт умением опытных наблюдателей смотреть краешком глаза, т.е. использовать периферическое (палочковое) зрение. Палочки, как известно, почти на два порядка более чувствительны, чем колбочки, обеспечивающие центральное зрение.
Для опытных испытуемых пороговая реакция на вспышки длительностью меньше 1 мс возникала в среднем при 17 квантах, падающих на роговицу, для менее опытных - 30. Величина порога зависела от длительности вспышки. Самые слабые вспышки фиксировались глазом только при малых длительностях - менее 1 мс. Реакция глаза на продолжительные световые стимулы (до 1 с) увеличивала пороговое значение до 200 квантов.
Харитон и Ли отметили одну интересную физиологическую особенность: длительное голодание и прием тонизирующих лекарств снижали порог с 17 до 12 квантов. Механизм такой сенсибилизации до сих пор не понят.
Установка С.И.Вавилова
Если Харитон решал задачу счета a-частиц, то Вавилов как специалист по оптике и люминесценции изучал флуктуации света. Глаз человека оставался незаменимым оптическим инструментом. Выступая на первой конференции по физиологической оптике, проходившей в Ленинграде в 1934 г., Вавилов говорил: “Исключительная чувствительность глаза в темноте и наличие резкого порога зрительного ощущения дают возможность визуально наблюдать флуктуации светового потока”. Еще в 1927 г. в первом издании книги “Глаз и Солнце” он писал:
“…мгновенно глаз в состоянии почувствовать очень небольшое число квантов. <…> Пользуясь этим, можно глазом обнаружить прерывистое, квантовое строение света <…>. Глаз, таким образом, действительно “воочию” позволяет убедиться в квантовой, прерывистой структуре света. Замечательно, что таким способом определяется не чувствительность глаза как целого, а чувствительность только последних клеток (палочек), ответственных за зрительное возбуждение. Отдельные кванты стали, в буквальном смысле слова, видимыми” [4].
Вавилов здесь подчеркнул важнейшее обстоятельство физиологии зрения: порог возникновения светового ощущения, т.е. работы зрительной системы в целом, и возбуждения отдельной фоторецепторной клетки - это вовсе не одно и то же, они существенно различаются.
Вавилов исходил из того, что при очень малом световом потоке (а это как раз вблизи порога зрительного ощущения) его флуктуации становятся значительными, поэтому именно они и ограничивают восприятие света. Действительно, при малом числе регистрируемых глазом фотонов n0 (от единиц до полутора-двух сотен) эти случайные числа подчиняются распределению Пуассона, для которого характерные флуктуации пропорциональны (n0)1/2. Уверенное выделение глазом слабой световой вспышки возможно, только когда n0 >> (n0)1/2 (сигнал превышает флуктуационный разброс), т.е. при достаточно больших n0. Относительно самой величины n0 - статистически среднего значения для каждого наблюдателя - Вавилов предположил, что в темноте порогу зрительного восприятия коротких световых вспышек соответствует какое-то минимальное, для каждого свое, число поглощаемых фотонов. По его словам, “единственный метод получить n0 дают лишь статистические измерения, вроде примененных нами. Таким образом <…> будет найдено действительное число фотонов, поглощаемое сетчаткой у порога раздражения. Все потери энергии, которые возникают в глазном яблоке вследствие отражения, поглощения и рассеяния и которые неизбежно влияют на любые энергетические методы, отпадают при статистических измерениях автоматически” [5].
С 1932 по 1941 г. в Государственном оптическом институте были выполнены сотни флуктуационных измерений с участием свыше десяти наблюдателей. Перед Вавиловым и его сотрудниками стояла задача добиться корректных условий опыта: кратковременности световых вспышек на фоне темновой адаптации, освещения небольших участков на периферии сетчатки при строгой фиксации их положения.
За 10 лет работы установка Вавилова несколько раз усовершенствовалась, но общая схема и основные элементы сохранялись [6]. Положение головы наблюдателя закреплялось с помощью подбородника таким образом, чтобы во время опыта глаз все время был фиксирован на красную сигнальную лампочку и свет от основной электрической лампы (видимый как зеленая точка) падал на периферию сетчатки - в 8° от ее центра. Свет от лампы проходил к глазу через диафрагму, зеленый фильтр, стопу стеклянных пластинок и нейтральный оптический клин. Между глазом и лампой располагался вращающийся диск с отверстием, размеры которого обеспечивали кратковременность световых вспышек (0.1 с). Для измерений с различными длинами волн вместо лампы использовали монохроматор с источником света. В ходе опыта интенсивность светового потока постепенно понижалась сначала реостатом, затем добавлением стеклянных пластинок и введенеием оптического клина. Наблюдатель отмечал каждую видимую вспышку нажимом ключа. По числу вспышек и прохождений света через отверстия диска, автоматически фиксируемых на бумажной ленте, определяли вероятность видения вспышек.

Схема второго варианта установки С.И.Вавилова. S - красная лампочка; G - стеклянная пластинка; L - лампочка, питаемая от аккумулятора; m - зеркало; O - диафрагма, закрытая молочным стеклом; F - зеленый фильтр; P - стопа стеклянных пластинок; K - нейтральный оптический клин; D - диск, насаженный на ось синхронного мотора (M); T - эталонный источник света (абсолютно черное тело).
Чтобы найти абсолютное число фотонов N, падающих на зрачок глаза в условиях порога восприятия, использовали эталонный источник энергии. Свет излучало нагретое тело, вплотную придвинутое к диафрагме (со снятым матовым стеклом). Зная геометрические параметры потока и характеристики излучателя как черного тела, экспериментаторы могли с достаточной точностью рассчитать (по закону Стефана-Больцмана) энергию фотонов, формирующих световое пятно на зрачке. Эти измерения в сине-зеленой области спектра для разных наблюдателей дали N = 108-335 фотонов, в среднем 208. Различие между n0 и N свидетельствовало о том, что большая часть энергии падающего на глаз света теряется на пути к фоторецепторам из-за отражения, поглощения и рассеяния в глазных средах и тканях.
Большинство измерений было сделано в сине-зеленой области спектра 500-550 нм. В этой области, как следовало из опытов Вавилова и его сотрудников, значения n0, соответствующие порогу на сетчатке, для одного и того же наблюдателя достаточно постоянны, но у разных людей могут быть различны. В работе 1933 г. пороговое число фотонов составило n0 = 47; в работе 1934 г. n0 = 8, а в сводной таблице (по результатам всех опытов в 1932-1941 гг.) n0 = 20.
Эксперименты С.Хехта
В начале 40-х годов американские физиологи С.Хехт, С.Шлер и М.Пирен опубликовали результаты своих исследований по определению абсолютного зрительного порога. Эта работа до сих пор признается классической. Определение минимального числа квантов проводили, измеряя минимальную энергию света, падающего на глаз, который вызывает “видение” вспышки. Установка Хехта отличалась от схемы Вавилова лишь некоторыми модификациями. Так, угол фиксации глаза на красную точку, который обеспечивал периферическое зрение, был бOльшим (20°), угловой размер основного пятна на сетчатке - тоже бо?льшим (10°). Свет (l= 510 нм) проходил к наблюдателю через нейтральный фильтр, оптический клин и двойной монохроматор. Длительность вспышки (1 мс) обеспечивал специальный затвор, а интенсивность света изменялась перемещением клина. Наблюдатель нажимом ключа производил вспышку (обычно около 50 вспышек одной и той же интенсивности) и сообщал, видел ли он ее или нет. Число фотонов, падающих на зрачок, пересчитывалось после измерения энергии света термоэлементом, установленным на диафрагме.

Схема установки С.Хехта. Г - глаз наблюдателя; P - диафрагма; FP - красная точка; FL - линза; D - диафрагма; L - лампа, питаемая от аккумулятора; F - нейтральный фильтр; W - оптический клин; М1М2 - двойной монохроматор; S - затвор. При нажиме кнопки затвор открывает путь световому потоку (l = 510 нм) на 1 мс.
Минимум энергии света на роговице, при котором наблюдатели (7 человек) фиксировали вспышки, варьировал в пределах (2.1-5.7)·10–10 эрг, соответственно чему и пороговое число фотонов сине-зеленого диапазона составило 54-148. Эти результаты перекрывались с данными Вавилова (108-335 фотонов), но были существенно больше, чем оценка Харитона и Ли (17 фотонов). По мнению Хехта, последние значения слишком малы [7].
Для уточнения пороговых значений непосредственно на сетчатке Хехт и соавторы учли, что 4% падающего на глаз света отражается от роговицы, почти 50% поглощается хрусталиком и по крайней мере почти 85 % оставшегося света проходит через сетчатку, не поглощаясь ею. Говоря другими словами, если весь свет, падающий на сетчатку, принять за 100%, то лишь около 15% поглощается ею, а остальные уходят в черный пигментный эпителий, лежащий за сетчаткой. После проведения такой корректировки получалось, что из 54-148 фотонов, падающих на роговицу, сетчаткой поглощается лишь 5-14. Оценивая размеры освещаемой области сетчатки (~500 палочек) и считая, что каждый фотон поглощается только одной палочкой, авторы пришли к выводу: вспышку можно “увидеть”, только если не менее 5-14 палочек почти одновременно поглотят по одному фотону.
Уточненная статистическая оценка флуктуаций потока, полученная Хехтом, давала для зрительного порога на сетчатке пять-восемь фотонов, что очень хорошо согласовалось с результатами измерений. Отметим здесь достаточную близость оценок Хехта (n0 = 5-8) и Вавилова (n0 = 20).
Современные представления
Итак, главный вывод из ранних работ Харитона и Ли (20-е годы), Вавилова и сотрудников (30-е годы) и Хехта и коллег (40-е годы) состоит в том, что зрительная клетка сетчатки - палочка - возбуждается при поглощении даже одного фотона. В ней фотон поглощается одной из 109 молекул зрительного пигмента - родопсина. Палочка должна каким-то образом “узнать” возбужденную молекулу и ответить на это одноквантовое событие возникновением электрического (рецепторного) сигнала. В последние годы удалось впрямую зарегистрировать этот очень слабый электрический сигнал [8]. В результате стало ясно: ответ зрительной клетки (и палочки, и колбочки) на единичный фотон есть событие дискретное, не зависит от интенсивности света, длительности вспышки и длины волны (цвета). У колбочек, однако, его величина оказалась слишком мала для того, чтобы возник такой рецепторный сигнал, который передавался бы следующим нейронам сетчатки. Этим объясняется относительно низкая (примерно на два порядка величины) чувствительность колбочек по сравнению с палочками.
В палочке электрический рецепторный ответ на поглощение одного фотона достаточно велик (около 3% от максимального значения), чтобы пройти через синапс к следующим нейронам сетчатки. Механизм трансдукции в палочке более эффективен, чем в колбочке. Под трансдукцией понимают преобразование энергии фотона, поглощенного молекулой родопсина, в электрическую энергию фоторецепторного сигнала. Ощущение же слабой световой вспышки возникает только при суммировании в нервных клетках сетчатки сигналов от нескольких возбужденных палочек, причем в течение небольшого промежутка времени.
Зрительная система человека способна распознавать как слабую вспышку одновременное поглощение 5-7 фотонов в рецептивном поле, насчитывающем около 500 палочек. Физический предел абсолютной световой чувствительности палочки определяется не только высокой эффективностью процесса трансдукции, но и низким уровнем биологического темнового шума.
Трансдукция запускается поглощением фотона хромофорной группой (11- цис ретиналем) в молекуле родопсина. Достаточно большая энергия, которую доставляет поглощенный фотон, тратится на фотоизомеризацию 11- цис ретиналя. Вместе с тем из-за тепловых флуктуаций молекула родопсина может с некоторой вероятностью активироваться и без поглощения фотона (так называемая реакция спонтанной темновой изомеризации). Такую темновую изомеризацию палочка воспринимает как ложный световой сигнал. Вероятность тепловых изомеризаций крайне мала: при комнатной температуре ее полупериод достигает 500-1000 лет. Это означает, что молекула родопсина исключительно стабильна. Казалось бы, темновой шум в зрительной системе должен быть ничтожен. Но в наружном сегменте палочки содержится огромное количество молекул родопсина (у черепахи и лягушки ~109, у человека ~108), и суммарное число спонтанных изомеризаций в каждой палочке может быть не столь уж мало. Поэтому в зрительной системе возникает шум, который человек, долго находясь в полной темноте, может “увидеть” как очень редкие случайные световые вспышки.
Итак, палочка способна уверенно детектировать один фотон, т.е. представляет собой эффективный счетчик квантов света. Сейчас достаточно ясен молекулярный механизм, обеспечивающий высокую чувствительность палочки [9]. Фотоизомеризация 11- цис ретиналя в молекуле родопсина запускает каскад ферментативных реакций, усиливающий сигнал в 105-106 раз. В результате в наружном сегменте палочки возникает электрический сигнал, который распространяется вдоль клетки по плазматической мембране и передается в первом синапсе следующим нервным клеткам сетчатки. По существу фототрансдукция - это вариант классической цепной реакции, подобной атомному взрыву, только происходящий в светочувствительном наружном сегменте зрительной клетки.
Внутри наружного сегмента палочки находится несколько сотен фоторецепторных дисков, каждый из которых содержит до 106 молекул зрительного пигмента родопсина. В мембране фоторецепторного диска находятся также основные белки - участники процесса трансдукции: Р - родопсин, Т - трансдуцин или G-белок и ФДЭ - фермент фосфодиэстераза; в цитоплазме наружного сегмента - фермент гуанилатциклаза - ГЦ. В темноте родопсин, трансдуцин и фосфодиэстераза неактивны. Ионный канал в плазматической мембране открыт благодаря “сидящим” на нем молекулам циклического гуанозинмонофосфата - цГМФ. Через открытый ионный канал внутрь клетки по градиенту концентрации поступают ионы натрия (Na+) и кальция (Ca++). В результате на мембране поддерживается электрический потенциал –40 мВ. При поглощении кванта света молекулой родопсина происходит изомеризация хромофора (11- цис ретиналя) и изменяется конформация белковой части молекулы. Это приводит к обесцвечиванию родопсина, который активирует трансдуцин (Т), в свою очередь взаимодействующий с фосфодиэстеразой (ФДЭ). Активированный ФДЭ гидролизует цГМФ, вследствие чего его концентрация в цитоплазме наружного сегмента падает. Ионный канал, который теряет цГМФ, закрывается, плазматическая мембрана гиперполяризуется, и возникает электрический нервный сигнал. Цепная реакция Р-Т-ФДЭ приводит к усилению светового сигнала в 105-106 раз.
Одно из удивительных и важных свойств палочки как счетчика одиночных фотонов - постоянство формы и величины электрического отклика, которое обеспечивается строго определенной геометрией клетки. Палочка напоминает фотоумножитель с сотнями или даже тысячами ориентированных фоторецепторных дисков. В плазматической мембране, окружающей всю клетку, равномерно распределены ионные каналы. Если световая вспышка достаточно яркая и насчитывает не десяток, а множество фотонов, они поглощаются молекулами родопсина по всей длине наружного сегмента. И тогда электрический ответ палочки существенно выше. Но все равно этот относительно большой электрический сигнал всего лишь сумма однофотонных сигналов.
Иными словами, элементарный акт в работе зрительной клетки - ее электрический ответ на поглощение единичного фотона. Таким образом, достаточно высокая энергия изомеризации 11- цис ретиналя, специфичность и высокая эффективность фототрансдукции, удивительное постоянство параметров фоторецепторного электрического ответа на поглощение единичного фотона и, возможно, некоторые другие механизмы и обеспечивают работу палочки в режиме счетчика фотонов с высоким отношением сигнал-шум.
Итак, абсолютная световая чувствительность зрительной системы (глаза и мозга) определяется наименьшим количеством световой энергии, которое вызывает субъективное ощущение света. В настоящее время порог светового восприятия экспериментально определен в (4-7)·10–10 эрг/с. Это - минимальный поток световой энергии от точечного источника, который падает на роговицу глаза и воспринимается мозгом как вспышка света. Для фотонов с длиной волны 507 нм (максимум кривой видности палочкового зрения) такая пороговая энергия соответствует 50-150 фотонам. Около половины этой энергии теряется на пути к сетчатке в оптических средах глаза, главным образом за счет отражения от роговицы и поглощения в хрусталике и стекловидном теле. Из доходящих до сетчатки 25-75 фотонов собственно фоторецепторными клетками поглощается всего 5-15, остальные (называемые “лишними”) проходят сквозь сетчатку и поглощаются лежащим за ней однослойным черным пигментным эпителием.
Природа феномена предельной световой чувствительности зрительной клетки находит свое объяснение. Заслуга С.Лэнгли, Ю.Б.Харитона, С.И.Вавилова, С.Хехта и многих других исследователей состоит в установлении самого этого феномена: одного поглощенного светового кванта достаточно для физиологического возбуждения рецептора сумеречного зрения - палочки сетчатки глаза.
Литература
1. Langley S.P. // Proc. Amer. Acad. Sci. 1881. V.16. P.342.
2. Langley S.P. // Phil. Mag. 1889. V.27. Series 5, 1.
3. Chariton J., Lea C.A. // Proc. Roy. Soc. 1929. V.CXXII.-A. P.304-352.
4. Вавилов С.И. Глаз и солнце. М., 1927.
5. Вавилов С.И. Флуктуации света и их измерения визуальным методом // Тр. физиол. оптики. Л., 1936. С.332-342.
6. Брумберг Е.М., Вавилов С.И. // Изв. АН СССР (ОМЕН). 1933. №7. С.919-941.
7. Hecht S., Shlaer S., Pirenne M.H. // J. General Physiology. 1942. V.25. P.819-840.
8. Островский М.А., Говардовский В.И. Механизмы фоторецепции позвоночных // Физиология зрения. М., 1992. С.5-59.
9. Pepe U.M. // J. Photochem. Photobiology. 1999. V.48. P.1-10.