Гипотеза Джанзена—Коннела
Одно из таких объяснений —гипотеза Джанзена—Коннела: ее в несколько разных терминах независимо сформулировали в 1970 г. два американских исследователя — Д.Джанзен и Ж.Коннел. Суть гипотезы сводится к тому, что в тропических лесах с очень высоким разнооб-разием деревьев семена и проростки имеют гораздо больше шансов выжить, если они находятся вдали от родительских растений. Джанзен полагал, что значительную часть семян, упавших рядом с «родителем»,поедают специализированные«хищники» (на самом деле растительноядные животные), которым гораздо легче разыскать свои излюбленные кормовые объекты, если они располагаются скоплениями. Коннел считал,что усиленное истребление более массовых видов деревьепроисходит чуть позже — на стадии проростков, и решающую роль играют листогрызущие насекомые, например, гусеницы тех или иных видов бабочек, кормящиеся определенными видами.Заметим, что и Джанзен и Коннел исходили из предположения о строгой пищевой специализации животных тропических лесов. Считалось, что у каждого растения есть свои потребители, которые в свою очередь контролируются специфическими хищниками и паразитами. Сразу заметим, что каких-либо серьезных эмпирических данных, подкрепляющих эти предположения, тогда небыло. Позднее же чешский энтомолог В.Новотный, работавший с большой группой помощников в тропическом лесу Новой Гвинеи, на основе огромного материала убедительно доказал, что подавляющее большинство листогрызущих насекомывовсе не узкие специалисты, а питаются по крайней мере не- сколькими видами растений [3,4]. Более того — средний уровень специализации насекомых в тропиках оказался не выше, чем у насекомых, обитающих в широколиственных лесах Центральной Европы.Надо сказать, что строгая проверка гипотезы Джанзена— Коннела в полевых условиях дело очень непростое. Результаты получались порой противореивыми, а кроме того, не прекращались споры о том, каковы должны быть размеры площадок для учета численности растений разного возраста. Однако проведенное недавно по единой схеме крупномасштабноизучение семи участков тропических лесов (два в Центральной Америке и пять в Юго-Восточной Азии) позволило сделать важный вывод: по мерроста и развития деревьев (от семян до зрелых растений) их разнообразие увеличивается,а позднее, на стадии старения и отмирания, снижается [5]. Разнообразие оценивалось как число видов с определенным числом особей, причем анализ проводился отдельно для четырех возрастных групп деревьев и на пробных площадках разной площади. Интервал между двумя обследованиями составил 5 лет, а для двух участков — 10 лет (с промежуточным обследованием через 5 лет).Как заметили авторы этой работы, обнаруженную ими лучшую выживаемость редких видов в сравнении с массовыми можно объяснить не только ги-
потезой Джанзена—Коннела. Не исключено, что эффект свя- зан с некоторым разделением экологических ниш, в результате чего интенсивность внутри- видовой конкуренции становится ниже межвидовой. Еще одна возможность — положительное влияние соседних растений, если они оказываютсдругого вида (это так называемая гипотеза облегчения)Но каковы бы ни были конкретные процессы, происходящие в сообществе, очевидно, что у редких видов вероятность выживания на ранних стадиях выше, чем у массовых.
Индексы разнообразия
Полученный эмпирически ряд убывания числа видов по мере возрастания их численности Фишер предложил описывать логарифмическими рядами. На основе такого статистического распре- деления разработали индекс разнообразия, количественно оценивающий скорость уменьшения числа видов в выборке помере увеличения числа особей.
Сообщество
сообщество — высоко интегрированная система, развивающаяся и реагирующая на изменения среды как нечто целое(порой напоминающее организм), или же это достаточно произвольнвыделяемая исследователями совокупность особей разных видов, волею судеб оказавшихся соседями, у которых особенности физиологии,строения и жизненного цикла позволяют существовать на определенной территории. Тот же вопрос можно сформулировать
несколько иначе: что важнее для поддержания структуры и функционирования сообщества —
система взаимосвязей между видами, жестко ограничивающая индивидуумы разных видов, или же свойства отдельных видов, позволяющие им выживать в ряду поколений вне зависимости от того, какие виды соседствуют с ними. Подчеркнем еще раз, что речь идет только о сообществах, образованных организмами одного трофического уровня (их иногда называют гильдиями). Мы не рассматриваем те, в которых продукты выделения одной группы организмов служат пищевыми ресурсами для другой (как это часто бывает в бактериальных системах,в частности в разных биопленках). Фактически это уже экосистемы, и принципы их орга
Информационный индекс!
Чуть позже Р. Маргалеф [Margalef, 1958], ссылаясь на формулу Л. Больцмана для энтропии изолированных термодинамических систем, предложил другое выражение для индекса разнообразия:
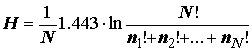 ,
,  (4.10)
(4.10)
где N! – факториальная величина всех исследуемых видов, ni! – факториал от числа особей каждого вида. Сопоставляя формулы (4.6) и (4.9), нетрудно увидеть, что (4.10) – просто иная форма расчета энтропии по Шеннону, не нашедшая, впрочем, широкого применения, поскольку, например, факториал от 34 составляет 3 1038 и является последним факториалом, вычисляемым на современном компьютере без использования специальных приемов.
Использование индекса Шеннона Н в экологии связано с рядом его действительных и мнимых свойств, которые нам представляется интересным обсудить:
“Разнообразие в индексе Шеннона трактуется как приходящееся на одну особь количество информации, заключенное в распределениях по видам, особям, или энергии по трофическим связям ” [MacArthur, 1955;цит. по: Одум, 1986]. Как уже было рассмотрено выше, энтропия Н отражает лишь один единственный аспект – степень выравненности вероятностей независимых событий (т.е. степень неопределенности встретить какой-либо вид). Сама по себе используемая оценка вероятности pi = Ni / N i полностью исключает учет в Н каких-либо данных об абсолютном количестве организмов, либо их биомассе. Одинаковые значения pi получаются как при больших (2000/10000), так и при малых (2/10) численностях особей, поэтому в интерпретациях индекса можно говорить только об относительном характере распределения информационных связей, вещества и энергии по отдельным таксонам, не затрагивая особь. И, наконец, сама по себе концепция независимости событий, заложенная в оценке энтропии, не предполагает оценку каких-либо структурных взаимодействий между таксонами, тем более трофических связей между ними.
“ Индекс Шеннона является мерой информации, содержащейся в экологической системе, подобно количеству информации в последовательности битовых сигналов в каналах связи” [Маргалеф, 1992]. Аналогия с каналами связи, где каждый очередной бит уменьшает неопределенность Н передаваемого сообщения и одновременно увеличивает количество принятой информации I, достаточно поверхностна, ибо никому еще не удалось разложить данные об экосистеме в последовательность взаимообусловленных квантов информации. К тому же, энтропия Н не вполне тождественна информации, а, с сугубо гносеологических позиций, является ее антиподом. И, наконец, информация о внутренней организации экосистемы, объективно присутствующая в структурах организмов, потоках между этими структурами, петлях обратной связи и проч., далеко не сводится только к разнообразию (тем более, к популяционной эквитабельности). Поэтому этот индекс, как заметил А.Ф. Алимов [2000], “скорее всего несет информационную нагрузку для исследователей, но не для экосистемы”.
“ Индекс Шеннона суммирует большое количество информации о численности и видовом составе организмов, учитывая число видов
R- И k-СТРАТЕГИИ
В 1967 г. Р. Макартур и Е. Уилсон, анализируя динамику численности
популяций, предложили r- и k - коэффициенты. Я не будем рассматривать их
математический смысл, а используем эти коэффициенты для обозначения двух
стратегий эволюционного развития живых существ. r-cтратегия предполагает
бурное размножение и короткую продолжительность жизни особей, а k-стратегия
- низкий темп размножения и долгую жизнь. В соответствии с r-стратегией
популяция развивается на переломных этапах своей истории, при изменении
внешней среды, что способствует появлению новых признаков и захвату новых
ареалов. k-стратегия характерна для благоденствия популяции при
сравнительно стабильных условиях. Очевидно, что у популяции вероятность
новаций будет тем выше, чем она быстрее размножается и чем чаще происходит
смена поколений, т.е. короче продолжительность жизни особей. Чтобы решить
проблему переходных форм, r-стратегии недостаточно, желательно дополнить ее
еще одним свойством, а именно повышенной жизнеспособностью, или лучшими
качествами в борьбе за существование, на коротком (в сравнении с k-
стратегией) временном промежутке, отведенном природой для жизни особи. Это,
в общем-то логично: за повышение жизнеспособности, как и за плодовитость,
приходится платить, а плата эта - сокращение продолжительности жизни.
Если жизнеспособность особей при r-стратегии повышена, это могло бы
компенсировать отмеченные недостатки промежуточных форм, связанные с
формированием новой функции. В результате они выстояли бы в борьбе за
существование.
Способность к переключению r- и k-стратегий есть один из механизмов
биологической эволюции. Чтобы остаться в пределах представлений об эволюции
как закреплении случайно возникших новых признаков путем естественного
отбора, нужно принять также, что и переключение стратегий происходит без
всякой закономерности, а выживают те, кто выбрал стратегию, более
соответствующую данным условиям среды. В простейшем случае должен
существовать какой-то один ген или координированная группа генов, режим
работы которых и определяет выбор стратегии.
Теория r-K отбора — в экологии теория, определяющая две различные природные стратегии размножения живых организмов, при определённых обстоятельствах обеспечивающие оптимальную численность для данного вида. Теория была разработана в 1967 году двумя американскими экологами — Робертом МакАртуром и Эдвардом Уилсоном.[1]
Общее представление
Согласно теории, естественный отбор в процессе эволюции происходит по одному из двух возможных сценариев, или стратегий. Эти стратегии, названные r и K, математически взаимосвязаны уравнением популяционной динамики Ферхюльста (англ. Verhulst equation):
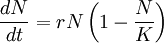
где r — скорость роста численности популяции (N), а K — перносимый объём, или максимальная плотность популяции. Проще говоря, организмы, придерживающиеся r-стратегии (так называемые «оппортунистические »), стремятся к максимально возможной скорости роста численности (параметр r). Потомство таких видов с большой долей вероятности не доживает до зрелого возраста. Организмы, придерживающиеся K-стратегии («равновесные »), наоборот находятся в состоянии равновесия со своими ресурсами и воспроизводят относительно мало, однако стремятся вложить в потомство как можно больше.
[править]r-K отбор и устойчивость к воздействиям внешней среды
В нестабильной или непредсказуемой окружающей среде преобладает r-стратегия, так как в этом случае ключевую роль играет способность быстрого размножения, а адаптационныемеханизмы, позволяющие конкурировать с другими организмами, ввиду быстро изменяющихся условий не столь важны. Характерными чертами r-стратегии являются высокая плодовитость, небольшие размеры, относительно короткое время жизни поколения и способность быстрого и широкого распространения. Среди организмов, выбравших r-стратегию, встречаются бактериии диатомовые водоросли, насекомые и сорные растения, а также головоногие и некоторые млекопитающие (особенно небольшие грызуны).
Если окружающая среда более-менее постоянная, то в ней преобладают организмы с K-стратегией, так как в этом случае на первое место выходит способность успешно конкурировать с другими организмами в условиях ограниченных ресурсов. Популяция K-стратегов, как правило, постоянна и близка к максимально возможной в данных условиях. Характерными чертами K-стратегии являются большие размеры, относительно долгий промежуток жизни и малое потомство, на воспитание которого отводится значительная часть времени. Типичными K-стратегами являются крупные животные — слоны, человекообразные обезьяны, бегемоты, киты и человек.
Сравнительный анализ обеих стратегий представлен в следующей таблице:[2]
| Характеристика | r-стратегия | K–стратегия |
| Численность популяции | Очень изменчива, может быть больше K | Обычно близка к K |
| Оптимальный тип местообитания или климата | Изменчивый и(или) непредсказуемый | Более-менее постоянный, предсказуемый |
| Смертность | Обычно катастрофическая | Небольшая |
| Размер популяции | Изменчивый во времени, неравновесный | Относительно постоянный, равновесный |
| Конкуренция | Часто слабая | Обычно острая |
| Онтогенетические особенности | Быстрое развитие, раннее размножение Небольшие размеры Единственное размножение Много потомков Короткая жизнь (менее 1 года) | Относительно медленное развитие Позднее размножение Крупные размеры Многократное размножение Мало потомков Долгая жизнь (больше 1 года) |
| Способность к расселению | Быстрое и широкое расселение | Медленное расселение |
[править]r-K как непрерывный спектр
Несмотря на то, что часть организмов являются исключительно r- или K-стратегами, большинство всё же имеют помежуточные характеристики между этими двумя крайними противоположностями. Например, у деревьев наблюдаются такие присущие K-стратегии черты, как долголетие и большая конкурентоспособность. Однако в процессе регенерации они вырабатывают тысячи плодов и распространяют их в большом диапазоне, что присуще r-стратегам.
2. Система Раменского–Грайма [38, с. 41–49; 37, с. 33–40; 55, с. 231–233; 77, с. 131–132]. Данный вариант типов стратегий отражает не только возможности выживания при разных затратах энергии на репродуктивное усилие, но и адаптации растений к различным условиям среды.
В качестве «ценобиотических типов» был предложен еще в 1935 г. отечественным ученым Л.Г. Раменским, который разделил все виды растений на виолентов (они же силовики, «львы»), патиентов (они же выносливцы, «верблюды») и эксплеренты (они же выполняющие, «шакалы»), отличающихся различными механизмами приспособления. Как видно из вторых названий, основными конкурентными преимуществами их были названы соответственно мощность (выражающаяся в захвате территории и удержании ее за собой, подавляя соперников энергией жизнедеятельности и полнотой использования ресурсов), выносливость (способность к выживанию в крайних, суровых условиях, постоянных или временных) и способность к быстрому захвату освободившихся вследствие изменения внешних условий экологических ниш.
Данная система осталась практически незамеченной даже в Советском Союзе, пока не была введена в научный оборот уже в конце 1970-х гг. биологом Т.А. Работновым. За границей о ней совершенно не знали. Одновременно с работами Работнова появились объемные монографии Дж. Грайма; последний повторил концепцию Раменского, что называется, «один к одному», завоевав широкий успех в научных кругах. Виоленты, патиенты и эксплеренты Раменского в его работах получили наименования конкурентов (C), стресс-толерантов (S) и рудералов (R).
В конце XX века данная классификация стратегий была не только переоткрыта, но и существенно детализирована. Так, Л. Раменский ограничился замечанием о том, что «в действительности большинство растений имеет характеристику переходную или смешанную, совмещающую черты двух типов; притом растения неодинаково ведут себя в различных местообитаниях». [55, с. 232]. Дж. Грайм же выделил 4 переходных типа стратегий, графически изображенных на рис. 1.3.

Рис. 1.3. Треугольник Грайма: C, R, S – первичные типы стратегий; CR, CS, RS, CRS – переходные (вторичные) типы стратегий
Признаки данных стратегий, характерные для различных представителей растительного мира, обобщены в табл. 1.3.
Судя по общебиологической литературе, значение перечисленных типов стратегий до настоящего времени еще полностью не осознано современными учеными данного направления. Разделение видов по K- и r-отбору используется главным образом в биологической экологии животных, классификация Раменского–Грайма – в экологии растений.
Таблица 1.3
Сравнительная характеристика типов стратегий растений на видовом уровне
[38, с. 45]
| Признак | Тип стратегии по Л. Раменскому | ||
| Виолент, силовик, «лев» | Патиент, выносливец, «верблюд» | Эксплерент, выполняющий, «шакал» | |
| Тип стратегии по Дж. Грайму | |||
| Конкурент (C) | Стресс-толерант (S) | Рудерал (R) | |
| Абиотические условия среды | Благоприятные | Неблагоприятные | Благоприятные |
| Наличие нарушений | Нет | Нет | Есть |
| Уровень конкуренции растений | Высокий | Низкий | Высокий |
| Жизненная форма | Деревья, кустарники, реже травы мезоморфного облика с широким простиранием в пространстве, мощной корневой системой и большой листовой поверхностью | Небольшие растения, кустарнички, деревья, многолетние травы ксероморфного (влаголюбивого – В.М.) облика, однолетние и многолетние суккуленты (выносливые к сухости, собирающие влагу – В.М.), лишайники, мхи | Однолетние травы, реже многолетние травы с интенсивным вегетативным размножением |
| Тип реагирования на стресс | Морфологический | Физиолого-биологический | Морфологический |
| Экологическая ниша | Широкая; по объему реализованная ниша близка к фундаментальной, дифференциация ниш выражена хорошо | Узкая; по объему реализованная ниша приближается к фундаментальной, дифференциация ниш не выражена | Широкая; по объему реализованная ниша много меньше фундаментальной, дифференциация ниш слабая |
В отличие от других положений биологии, связанных с адаптивными процессами и конкурентной борьбой, вторая классификация получила признание среди ученых-экономистов и после некоторой доработки получила распространение в отечественной экономической литературе (см. подразделы 2.3.3–2.3.4 настоящей работы).
Биоразнообра́зие (биологи́ческое разнообра́зие) — разнообразие жизни во всех ее проявлениях. В более узком смысле, под биоразнообразием понимают разнообразие на трех уровнях организации: генетическое разнообразие (разнообразие генов и их вариантов — аллелей), разнообразие видов в экосистемах и, наконец, разнообразие самих экосистем.А. В. Марковым и А. В. Коротаевым была показана применимость гиперболических моделей положительной обратной связи для математического описания макродинамики биологического разнообразия [1][2].
Признаки биоразнообразия
Биологическое разнообразие видов характеризуется двумя признаками — видовым богатством и выравненностью. Видовое богатство отражает число видов, встречающихся в пределах экосистемы, в то время как выровненность характеризует равномерность распределения численности животных[4]. Выделение этих компонентов связано с тем, что за редким исключением в экосистемах среди организмов, принадлежащих к одному трофическому уровню, экологической или таксономической группе, большая часть биомассы достигается за счёт вклада очень немногих видов. Биоразнообразие — ключевое понятие в природоохранном дискурсе. Каким-то объективным способом определить необходимость сохранения и поддержания биоразнообразия довольно трудно, поскольку это зависит от точки зрения того, кто оценивает эту необходимость. Тем не менее, существует три главные причины сохранять биоразнообразие:
С утилитарной точки зрения элементы биоразнообразия являются ресурсами, которые уже сегодня представляют реальную пользу для человека или могут оказаться полезными в будущем. Биоразнообразие как таковое приносит как экономическую, так и научную пользу (например, в поисках новых лекарственных препаратов или способов лечения). Выбор в пользу сохранения биоразнообразия — это этический выбор. Человечество в целом — это часть экологической системы планеты, и потому оно должно бережно относится к биосфере (в сущности мы все зависим от ее благополучия). Значимость биоразнообразия можно также характеризовать в эстетическом, сущностном и этическом плане. Природа прославляется и воспевается художниками, поэтами и музыкантами всего мира; для человека природа является вечной и непреходящей ценностью.
Разнообразия
В современной фауне насчитывается примерно 4000 видов млекопитающих и 9040 видов птиц. Описано около 19 000 видов современных рыб. Число всех известных ныне живущих видов позвоночных достигает примерно 42 000.
В некоторых других крупных группах число известных современных видов гораздо выше — описано 107 000 видов моллюсков, 215 000 видов цветковых растений и 750 000 видов насекомых. Оценки числа видов в разных крупных группах организмов приведены в табл. 1.1 и 1.2. Как видно из этих таблиц, общее число известных видов живых существ в современном мире составляет примерно 1.4 млн.
| Таблица 1.1. Примерное число описанных современных видов в царстве животных (по Mayr, 1969; Wilson, 1988; Mayr, 1988*) | ||
| Хордовые | 43 870 | |
| Млекопитающие | ||
| Птицы | ||
| Рептилии | ||
| Амфибии | ||
| Костные рыбы | 18 150 | |
| Акулы и скаты | ||
| Бесчелюстные | ||
| Оболочники и ланцетники | ||
| Членистоногие | 837 000 | |
| Насекомые | 750 000 | |
| Ракообразные, паукообразные и т. п. | 123 000 | |
| Молюски | 107 250 | |
| Брюхоногие | 80 000 | |
| Двустворчатые | 25 000 | |
| Иглокожие | ||
| Кольчатые черви | 12 000 | |
| Плоские черви | 12 200 | |
| Нематоды и родственные им группы | 12 500 | |
| Кишечнополостные | ||
| Мшанки и родственные им группы | ||
| Губки | ||
| Разные мелкие группы | ||
| Всего низших беспозвоночных | 69 700 | |
| Общее число видов в царстве животных | 1 093 820 |
Сложнейшая задача таксономического изучения и описания довольно хорошо решена для таких групп, как птицы и млекопитающие, но далека от завершения для большинства остальных групп. Значительное число морских беспозвоночных, цветковых растений и представителей других групп всё ещё ждет своего таксономического описания. По оценкам ихтиологов, общее число видов современных рыб приближается к 40 000, из которых пока описано лишь 20 000. Объём работы, которую предстоит проделать систематикам насекомых, ещё больше. По мнению энтомологов, описанные до сих пор виды насекомых (около 750 000) составляют лишь небольшую долю, возможно одну десятую, всех видов насекомых, действительно обитающих на Земле в настоящее время.
| Таблица 1.2. Примерное число современных описанных видов в четырёх царствах | ||
| Царства и основные группы | Примерное число видов | источник данных* |
| Царство растений | ||
| Цветковые растения | 215 000 | Cronquist, 1981 |
| Голосеменные растения | Jones, 1941 | |
| Папоротники и близкие к ним группы | 10 000 | » » |
| Мхи и печёночники | 23 000 | » » |
| Зелёные водоросли | 7 500 | Bold et al., 1980 |
| Бурые, красные и золотистые водоросли | 11 400 | » » |
| Всего | 267 540 | |
| Царство грибов | ||
| Настоящие грибы | 40 000 | Ainsworth, Bisby, 1954 |
| Миксомицеты | 4 000 | » » |
| Всего | 44 000 | |
| Царство протистов | ||
| Простейшие | 30 800 | Wilson, 1988 |
| Эвгленовые | Bold et al., 1980 | |
| Динофлагеллаты | ~1 000 | Bold (личное сообщение) |
| Всего | 32 600 | |
| Царство монер | ||
| Синезелёные водоросли | 1 500 | Bold et al., 1980 |
| Бактерии | 3 060 | |
| Всего | 4 560 | |
| Общее число для 5 царств | 1 442 520 |
Если принять довольно скромные допущения о соотношении известных и неизвестных видов в различных крупных группах, то можно получить приблизительные оценки общего разнообразия существующих видав. Число ныне живущих видов составляет не менее 5., а возможно и 30 млн. (Grant, 1963; Wilson, 1988*). Были сделаны две независимые оценки числа видов как ныне живущих, так и вымерших, существовавших на Земле за всю её геологическую историю (Simpson, 1952; Grant, 1963*). Диапазоны этих оценок в значительной мере перекрываются. С учетом этого можно принять, что общее число видов за всю историю жизни на Земле составляет от одного до нескольких миллиардов.
На Земле существует (по разным данным) от 1,2 до 2 млн. видов животных и растений. Из них на долю растений приходится примерно 1/4 или 1/3 общего числа видов. Из животных по числу описанных видов первое место занимают насекомые (около 740 000), второе — моллюски (по разным данным, от 40 000 до 100 000), затем идут позвоночные (60 000 — 70 000 видов). Из растений на первом месте — покрытосеменные (по разным данным, от 150 000 до 290 000 видов), затем грибы (от 70 000 до 100 000 видов). Числом видов растений и животных измеряется богатство флоры и фауны.
Однако обилие видов ещё не означает обилия особей, так же как и бедность флоры и фауны видами может сопровождаться чрезвычайным обилием особей. Поэтому для характеристики растительности и животного мира, в отличие от флоры и фауны, пользуются понятиями биомассы (общей массы организмов) и биологической продуктивности — способности организмов к воспроизводству биомассы в единицу времени (на единицу площади или объёма местообитания). По биомассе организмы распределяются иначе, чем по числу видов: биомасса растений на суше значительно больше, чем животных.
Насекомы в тропиках
В тропических лесах термиты являются основными разрушителями всех растительных остатков. Образование почвы в тропиках и перемешивание ее слоев в значительной степени зависят от деятельности термитов [13].
Важнейшая группа тропических насекомых — термиты. Они непрерывно разрушают и рассеивают в почве в виде своих экскрементов остатки древесины и старые, опавшие на землю листья деревьев и лиан, подготавливая таким образом растительные вещества к минерализации бактериями и грибами. Без достаточного количества термитов тропические гилей не могут нормально существовать: их почва становится малоплодородной, уплотняется, начинают гибнуть прорастающие из семян и уже развивающиеся молодые растения.
Вторая важнейшая группа тропических насекомых — живущие в дуплах, в скалах и открыто на ветвях деревьев общественные пчелы, родственные нашим домашним. Дикие тропические пчелы нередко более ярко окрашены, крупнее или же, наоборот, мельче домашних. Поскольку среди густой растительности гилей при очень частых дождях и в насыщенном влагой воздухе перекрестное опыление растений при помощи ветра почти невозможно, пчелы оказываются главными опылителями подавляющего большинства тропических лесных растений. В опылении растений участвуют также некоторые бабочки, мухи и мелкие птицы — колибри в тропиках Западного полушария и нектарницы в Восточном полушарии.
И третья совершенно необходимая группа животных гилей, опять-таки из класса насекомых, — это муравьи. Десятки их видов и многие миллионы особей круглый год обшаривают деревья, лианы и поверхность почвы, нападая преимущественно на растительноядных насекомых: без этой защиты растения гилей, их листья и цветы очень быстро бы погибли.
Некоторые группы насекомых быстро уничтожают мертвые стволы деревьев, поддерживая круговорот веществ в природе. В тропиках практически единственными насекомыми-почвообразователями являются влаголюбивые термиты, перерабатывающие в лесах всю отмирающую древесину и растительные остатки. Кроме того, термиты принизывают почву своими многочисленными ходами и перемещают ее в различных направлениях, перемешивая слои. Пропуская растительную пищу и почву через кишечник, они активно влияют на происходящие в почве процессы. Сами же термиты представляют собой достаточно калорийный источник питания для многих животных. Несмотря на то что с деятельностью термитов сталкиваются интересы человека (насекомые разрушают деревянные постройки людей), роль термитов в жизни тропической природы, несомненно, огромна.
Фитофаги (от фито... и ...фаг), животные, пищей которых служат только растения. К облигатным (обязательным) Фитофаги относятся многие насекомые, среди которых наряду с поли- и олигофагами есть и монофаги (см. Монофагия). Многие растительноядные позвоночные (например, грызуны, «зерноядные» птицы) поедают и животную пищу, главным образом беспозвоночных.
Видоспецифичность (species-specifity) - Свойство какого-либо признака (всегда так или иначе генетически детерминированного) характеризовать только какой-то один вид организмов по сравнению с другими видами.
Распределение числа видов по размерным классам организмов
Пределах одной таксономической группы организмов виды разного размера встречаются с разной частотой. Для наземных млекопитающих распределение характеризуется крайней асимметрией: модальный класс приходится на довольно мелкие (но не самые мелкие) виды; видов с меньшей массой мало, а крупных и очень крупных — довольно много. С помощью простой модели удалось показать, что такое распределение может быть обусловлено всего несколькими достаточно универсальными факторами.
Если все виды млекопитающих разбить на размерные группы (по весу взрослых особей) и построить график зависимости числа видов от массы тела, то получится резко ассиметричное распределение. Модальный класс (наибольшее число видов) приходится на вес 40 г. При дальнейшем уменьшении массы тела число видов падает быстро, а при увеличении — очень медленно. Таким образом формируется асимметрия распределения — «хвост» из крупных видов, который растягивается далеко вправо — вплоть до африканского слона (Loxodonta africana), самого крупного из ныне живущих наземных млекопитающих (вес самца 5–7 т).
Наблюдаемая картина выявляется при сведении воедино данных, имеющихся для 4002 видов наземных млекопитающих — современных, а также вымерших сравнительно недавно, в конце четвертичного периода (рис. 1).
|
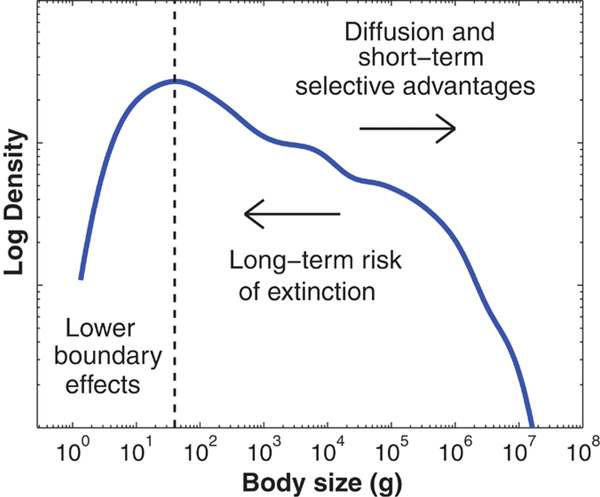 Рис. 1. Распределение массы тела для 4002 «рецентных» (современных и вымерших в позднем четвертичном периоде) видов сухопутных млекопитающих. По оси X — масса тела в граммах. По оси Y — относительное число видов (обе шкалы логарифмические). Модальный класс приходится на зверей с массой тела около 40 г, но, как хорошо видно из графика, крупных млекопитающих гораздо больше, чем мелких. Стрелка, обращенная вправо, — тенденция к увеличению размера тела, обусловленная кратковременным преимуществом более крупных организмов. Стрелка, обращенная влево, — тенденция к уменьшению размера, обусловленная высоким риском вымирания крупных форм. В левой части графика вступают в действие ограничения минимального размера (связанные главным образом с энергетикой). Рис. из обсуждаемой статьи в Science
Рис. 1. Распределение массы тела для 4002 «рецентных» (современных и вымерших в позднем четвертичном периоде) видов сухопутных млекопитающих. По оси X — масса тела в граммах. По оси Y — относительное число видов (обе шкалы логарифмические). Модальный класс приходится на зверей с массой тела около 40 г, но, как хорошо видно из графика, крупных млекопитающих гораздо больше, чем мелких. Стрелка, обращенная вправо, — тенденция к увеличению размера тела, обусловленная кратковременным преимуществом более крупных организмов. Стрелка, обращенная влево, — тенденция к уменьшению размера, обусловленная высоким риском вымирания крупных форм. В левой части графика вступают в действие ограничения минимального размера (связанные главным образом с энергетикой). Рис. из обсуждаемой статьи в Science
Какие факторы определяют подобное распределение размеров тела по всему классу, как соотносятся в эволюции тенденция к увеличению массы тела и отрицательные последствия крупного размера. На эти вопросы попытались ответить Аарон Клозе (Aaron Clauset) и Дуглас Эрвин (Douglas H. Erwin) из Института Санта-Фе (Нью-Мексико, США), предложившие модель эволюции размеров тела млекопитающих, которая исходит их трех предположений:
1) Изменение размера тела в ходе эволюции рассматривается как «кладогенетический мультипликативный диффузионный процесс» (то есть потомок наследует размер от предка с некоторыми случайными вариациями, но с тенденцией к увеличению). Возрастание массы тела в эволюционно прослеживаемой линии животных, так называемое правило Копа (Cope's rule), — эмпирическая закономерность, выявленная палеонтологами для некоторых (отнюдь не всех) групп животных. Считается, что более крупный размер тела повышает шанс противостоять хищникам, позволяет легче перенести голод и обеспечивает лучшую терморегуляцию. Кроме того, для более крупных животных меньше траты энергии на единицу массы.
2) Минимальный размер млекопитающих жестко ограничен. Самые мелкие известные млекопитающие имеют массу тела около 2 г — это один из видов землероек, карликовая белозубка (Suncus etruscus), и летучая мышь-шмель, или, как ее иначе называют, свиноносая летучая мышь (Craseonycteris thonglongyai). Интенсивность удельного (в расчете на единицу массы) обмена столь мелких зверьков чрезвычайно высока. Скорее всего, именно это обстоятельство является решающим в определении нижнего размерного предела млекопитающих.
3) Любой вид подвержен риску вымирания, причем вероятность его с увеличением размера тела возрастает. Уязвимость крупных видов к неблагоприятным воздействиям — следствие низкой плотности их популяций. Если численность популяций очень мала, то существует довольно высокая вероятность ее вымирания от какой-либо случайной комбинации неблагоприятных условий.
Гетеробатмия (от гетеро... и греч. bathmós — степень, ступень), неодинаковый уровень специализации различных органов, достигнутый в процессе эволюции организмов.
Мозаичная эволюция, одна из форм эволюции организмов, при которой органы и части тела развиваются независимо и неравномерно: одни быстрее, другие медленнее, третьи остаются практически неизменными длительное время. Термин «Мозаичная эволюция » предложен английским учёным Г. де Беером (1954). Результат Мозаичная эволюция — неодинаковый уровень развития и специализации разных о