Дифференсационные деревья, мусорное ДНК молекулярных часов, и
Эволюция неотении у саламандр.
(Кристофр Мартин и Ричард Гордон)
Отделения радиологии и ботаники * Университет Манитобы, Виннипег, R3t 2N2, Канада
Ключевые слова: Неотения; мусорная ДНК; С-значение; молекулярные часы; хвостатые.
Конспект
Облигатно неотеничные саламандры погибают, если форсируют метаморфоз. Мы предполагаем, что это можно объяснить, предположив, что; 1) их "избыточная" ДНК" является "мусорной "ДНК; 2) «взрослая» определяющая часть ДНК становится "мусорной" ДНК, но доступна для повторной дупликации. Это говорит о возможностях использования этой новооткрытой "мусорной" ДНК в концепции молекулярных часов. Мы получаем замечательное согласие в "предсказании" количества ДНК в ядре для на сегодняшний день необлигатно-неотенических саламандр из этих молекулярных часов. Эти наблюдения согласуются с идеей, что развитие этих животных вполне описуемо в терминах дифференсационных деревьев, ветви которых (генные каскады), соответствующие взрослым соматическим тканям, накапливают вредные мутации в течение эволюционного времени. Мы показываем, что количество ДНК на ядро линейно растёт с филогенетическим возрастом семейств саламандр. Недостаток сдерживания в естественном отборе на неиспользуемые взрослые ветви можно объяснить большим количеством так называемой" мусорной ДНК " у облигатно неотенических саламандрах. Эффекты этой избыточной ДНК, благодаря усилению клеточного размера, предполагают позитивную обратную связь, экофизиологическое объяснение для такой мусорной ДНК: адаптация к условиям холодной воды, усиливается более низким метаболизмом, связанным с бОльшим количеством ДНК, более крупными клетками и более растянутым временем развития.
Введение
Одной из главных нерешенных проблем эволюционной биологии является взаимосвязь между развитием индивидуального организма и его потенциалом для дальнейшей будущей эволюции (Боннер, 1965, 1982; Остер и Альберх, 1982; Хордер, 1983; Рафф и Кауфман, 1983; Артур, 1988).
Мы предложили новую теорию развития тканей у эукариотных организмов (Гордон, 1993), коренящуюся в биомеханической модели для дифференциации (Гордон и Brodiand, 1987; Бьерклунд и Гордон, 1993, 1994) на основе ультраструктуральных наблюдений (Бернсайд, 1971, 1973; Мартин и Гордон. 1995) и ограниченных временем наблюдений волн дифференциации (Бродянд и др., 1994; Gordon, 1995; Gordon et al., 1994; cf. Готов, Хэнсон и Бенцер. 1976) во время первичной нейронной индукции. Основная идея заключается в том, что дифференцировку можно рассматривать как временное чередование механизмов разрешения внутриклеточной физической нестабильности и каскада генов клеток определённого типа. Каждая ткань разделена в 2 новых ткани, клетки которой после этого раскрывают различные комплекты генов и генетических продуктов. Бифуркация тканей может быть представлена бинарным деревом, которое мы называем деревом дифференцировки организма (Фиг. 1). Поэтому дерево дифференцировки является деревом происхождения ткани. Последняя концепция восходит к фон Баеру (1828, см. Patterson, 1983). Slack (застой) (1983) приносит дерево происхождения тканей для земноводных.
Существует множество проверяемых разветвлений теории развития, основанной на дифференсационных деревьях (Gordon, 1995). Мы покажем, как дифференсационные деревья могут обеспечить разумную основу для организации известных фактов об эволюции неотенических саламандр. Поскольку факультативная и облигатная неотения, по-видимому, являются реакцией на местную окружающую среду (обзор ниже), эта статья демонстрирует первую из возможно существующих многих связяей между дифференсационными деревьями, экологией и эволюцией. В частности, мы находим, что количество ДНК на ядро линейно увеличивается с геологическим временем у облигатно неотенических саламандр, что даёт новое "мусорное" ДНК молекулярных часов.
Фиг.1 Дифференсационное дерево метаморфизирующих животных
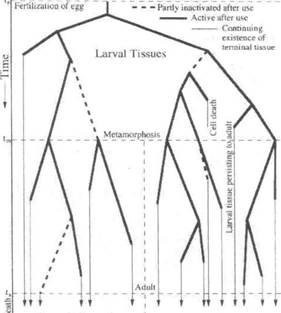
Половые клетки и Репродуктивные Ткани /Соматические Взрослые Ткани
Фиг1. Дифференсационное дерево метаморфизирующих животных. Время развития указано сверху вниз. Организм начинает как оплодотворенная яйцеклетка во время L0. Каждая толстая линия представляет собой генный каскад, его продукты, и морфогенетические изменения и клеточную форму этих изменений и т. д.. из всех клеток в одной, смежной ткани. Пунктирные толстые линии представляют собой генные каскады, которые частично или полностью отключаются, когда клетки проходят последующие этапы дифференцировки. Узлы представляют синхронное сооружение расхождений клеточного строения в каждой клетке в ткани. Каждая клетка в ткани разрешает нестабильность своего штатного клеточного расхождения путем подбора одного из 2 новых клеточных типов. Во время метаморфоза tm половые клетки, репродуктивные ткани и взрослые соматические ткани дополнительно дифференцируются от некоторых из личиночных тканей. Дифференцировка во взрослое состояние завершается в момент времени ta. Прямые линии, нисходящие от каждого терминального узла, указывают на продолжение существования окончательно дифференцированных клеточных типов, если только не происходит "запрограммированная" клеточная смерть (апоптоз).
Обзор неотении
Феномен неотении определяется как достижение репродуктивной зрелости при сохранении личиночной внешней морфологии и личиночной репродукции через замедленное соматическое развитие (Пирс и Смит, 1979). Андерсон и Уортингтон (1971) и Нуссбаум (1976) подтверждают это определение, показывая, что личинки Ambvsioma ordmarium и Dikamptodon ensatus достигают половой зрелости примерно в том же размере, что и прошедшие метаморфоз их сородичи. Предполагается, что связь между размером и возрастом остается такой же и у остальных метаморфизирующих и неотенических саламандр.
Метаморфоз
Прежде чем пытаться обсуждать неотению у саламандр, мы должны сначала понять нормальное состояние метаморфоза. Метаморфоз - это серия резких структурных, физиологических, биохимических и поведенческих изменений (Duellman и Trueb 1986) с участием:
1) регрессии структур и функций, которые характерны только для личинки;
2) трансформации личиночных структур в форму, подходящую для взрослых;
3) развития новых структур и функций, жизненно необходимых взрослым.
Метаморфоз влияет почти на каждую ткань в организме. В таблице 1 показаны основные морфологические изменения. Интересно отметить, что репродуктивные ткани (зародышевая линия и репродуктивные органы) не входят в число тех, которым сообщаются изменения при трансформации (Таблица 7.2 в Duellman и Trueb, 1986), что подтверждает наше предположение о том, что они образуются независимыми ветвями дифференсационного дерева (Фиг. 1). Прямое подтверждение их недостаточного изменения на трансформации стоило бы искать.
табл.1 (она же 7.2) Некоторые морфологические изменения, происходящие при метаморфозе саламандры
Образование кожных желез
Дифференцирование Leydig клеток
Увеличение retinal rodopsin
Увеличение белка в крови
Развитие легких (кроме Плетодонтидов)
Дегенерация складки хвостового плавника
Изменение пигментации
Дифференсация гемоглобина
Жаберная резорбция
Изменения почек
Метаморфоз контролируется эндокринной системой через влияние тиреоидных гормонов на ткани-мишени. Пролактин может сыграть ключевую роль в метаморфозе путем стимуляции периферийного преобразования Т4 к более активной форме Т3 (Андерсон и Дент, 1982). Однако, пролактин также хорошо подавляет работу тироидных гормонов на дегенерацию хвоста и резорбцию жабр. Хотя действие пролактина на неотению хвостатых амфибий не столь хорошо изучено, как в случае метаморфоза у бесхвостых. Пролактин действует как ювенильный или антиметаморфный гормон, препятствующий или замедляющий запрограммированную клеточную смерть во время развития личинок бесхвостых амфибий (Etkin и Gona, 1967). Тироидный гормон включает регрессию хвоста в культуре Xenopus и формирование участков почек в культуре задерживают аппликацию пролактина в средней культуре (Tata et al. 1991). Инъекция пролактина головастикам также повлияла на заметное уменьшение активности некоторых гидролитических энзимов найденных в лизосомах и растворимых фракциях (частично отвечающих за регрессию ткани) которые индуцировались тироидным гормоном (Blatt et a!., 1969; Дерби, 1975). Tata (1984) и Tata et al. (1991) предполагали, что пролактин может воспрепятствовать синтезу или функциональной активации предполагаемым гормонально-наведённым тироидным протеином или mesenchymal-ными регрессионными факторами. Доказательство также поддерживает концепцию что пролактин может подействовать на регуляцию экспрессию и функцию тироидного рецепторного гормона.
Неотения
На сегодняшний день выяснено, что метаморфоз не происходят у неотенических саламандр по двум причинам:
1) Линн (1961) показал, что клетки Proteus anguinus и Necturus spp. нечувствительны к гормонам щитовидной железы (тироидным).
2) Гипофиз у Amhystoma mexicanum видимо, нечувствителен к тироидному освобождающему гормону (TRH); поэтому он не в состоянии реализовать тироидный стимулирующий гормон (TSH), который включает в организме выделение тироксина из щитовидной железы (Jacobs с соавт.. 1988; Taurog et al., 1974).
Факультативная неотения
Для того, чтобы эволюционное значение неотении было понятным, должно быть установлено, должна ли являться неотения генетически контролируемой и генетически изменяемой для неотении, случающейся в популяциях саламандр. Если неотения контролируется исключительно окружающей средой, эволюционный потенциал первой ограничен. Взяв две разных субпопуляции личинок Ambvstoma talpoideum и выращивания их в общием искусственном пруду, Semlitsch и Gibbons (1985) показали различия в частоте неотении. Это свидетельствует о том, что факультативная неотения может иметь генетическую основу. Эксперименты Semlitsch и Wilbur (1989) показали, что наследуемая генетическая изменчивость существует, и предположили, что эволюция и дивергенция между популяциями саламандр возможны при интенсивной селекции за сравнительно короткие периоды экологического времени. Семлич и Вильбур (1989) искусственно отбирали для неотении в субпопуляциях факультативно неотнических A. talpoideum, которые были подняты в искусственный пруд. После четырех поколений искусственной селекции они наблюдали статистически значимое увеличение неотеников на 1,5%. Healy (1970) наблюдал естественную популяцию Notophthalmus viridescens в течение пятилетней засухи. Эта засуха коренным образом изменила экологические условия пруда, включая уменьшение плавающей и погруженной в воду растительности, снижение уровня воды и повышение температуры воды. Во время засухи процент неотеников значительно снизился. Результаты естественного или искусственного отбора для неотении, таким образом, очевидны через очень короткое время.
Томпкинс (1978) предположил, что неотения есть результат от гомозиготности по одному рецессивному гену, в то время как Харрис (1987) предположил, что неотения является полигенетически смешанным признаком, в результате сильного давления окружающей среды и условиях постоянных колебаний в численности популяции. В настоящее время истинная генетическая основа неотении остаётся неизвестной.
Высокий уровень встречаемости неотении происходит при различных условиях окружающей среды. Харрис (1987) показал, что неотения зависит от плотности популяции: высокая встречаемость неотеников наблюдается при низкой плотности личинок. Sprules (1974a, б) показал, неотении способствуют постоянные, прохладные водные локалитеты при суровых наземных условиях. Figel и Semlitsch (1990) коррелируют неотению с постоянными водными локалитетами без рыбы. Популяции Ambistoma gracile неотеничны на больших высотах, тогда как метаморфоз наиболее распространён в низинных популяциях (Снайдер, 1956). Данн (1931) предположил, что случаи неотении в горных районах могут быть вызваны низкими концентрациями йода, препятствующими производству тироксина (в щитовидной железе). Это никогда не было доказано экспериментально. Такие виды, как Proteus, обитающие в подземных водах пещер, исключительно неотеничны (Duellman and Trueb, 1986). Подземные климатические условия здесь заключаются в постоянной круглогодичной температуре 5 -10 С, относительно постоянном запасе пресноводных ракообразных и очень малом количестве хищников по отношению к суровым условиям наземной среды (Steward, 1969). Из вышеизложенного можно сделать вывод, что K-селективные среды благоприятствуют неотении. Неотеники не колонизируют новые ареалы и не пытаются удержаться в более суровых (в частности, сухопутных) условиях, используя R-селективные среды для быстрого воспроизводства. Однако они образуют стабильные популяции в благоприятных водоемах. Когда окружающая среда становится R-селективной, предпочтение отдается трансформации (Gould, 1977). Эти пункты очень поэтично описаны Гарстангом (Garstang, 1966):
"...Но когда подходяще озеро - аэрируется, обильно едой,
Они цепляются за вечную молодость и выводят личинок рой."
"But when a lake's attractive, nicely aired, and full of food
They cling to youth perpetual and rear a tadpole brood."
Облигатная неотения
Считается, что облигатные неотеники, такие как протеи и амфиумы, могли стать неотеническими в меловой период в ответ на суровые наземные условия (т. е. локальная водная среда относительно более стабильна), тогда как аксолотли факультативно неотенических Amhystoma spp. как полагают, имеют более недавнее происхождение, в Плейстоцене, при несколько иных климатических условиях (Dueliman и Trueb, 1986). Таким образом, продолжительность неотенической экспрессии может оказывать решающее влияние на степень выраженности неотении (облигатную или факультативную).
Одна замечательная характеристика облигатно неотенических саламандр заключается в том, что они, среди всех животных дублируют количество собственного ДНК в ядре:
"Большинство саламандр имеют 30-86 хромосом [ДНК] на ядро, но члены облигатно неотенических семейств имеют 91 -192 хромосом в расчете на одно ядро, уступая только лёгочным рыбам (lungfish; 160-284 хромосом на одно ядро). Однако, кажется, что нет никакого филогенетического значения в количестве ядерной ДНК внутри неотенических семейств саламандр или между саламандрами и двоякодышащими. Вместо этого существует тесная связь между увеличением количества ядерной ДНК и большим размером клеток в обеих группах (Thomson, 1972; Morescalchi, 1975)" (Dueliman and Trueb, 1986).