Теоретическая часть
Молекулярные основы наследственности.
Свойство организмов передавать потомству свои признаки и особенности развития, основано на передаче из поколения в поколение наследственной информации. В середине XIX в. было установлено, что способность к наследованию признаков определяется материалом, находящимся в ядре клетки. Саттон и Т. Бовери предположили, что хромосомы являются носителями наследственной информации. В 1869 г. швейцарский химик Ф. Мишер обнаружил в клеточном ядре особое вещество кислого характера, названное им нуклеином. Это событие расценивается как открытие нуклеиновых кислот. Потребовалось несколько десятков лет, чтобы убедиться, что материальным носителем наследственной информации является только одна из составных частей хромосомы – молекула дезоксирибонуклеиновой кислоты (ДНК).
Генетический код.
Генетический код – свойственная живым организмам единая система записи наследственной информации в молекулах нуклеиновых кислот в виде последовательности нуклеотидов; определяет последовательность включения аминокислот в синтезирующуюся полипептидную цепь в соответствии с последовательностью нуклеотидов ДНК гена (или процесс перевода триплетной последовательности нуклеотидов молекулы ДНК в последовательность аминокислот в белковой молекуле). В узком смысле генетический код – это словарь кодонов (триплетов иРНК), кодирующих те или иные аминокислоты и знаки пунктуации белкового синтеза.
Первичное действие гена. Аллельные состояния гена.
Гены, расположенные в идентичных локусах гомологичных хромосом, называются аллельными генами. Основными типами их взаимодействия являются:
|
|
— доминирование: если функциональное состояние одного аллельного гена не зависит от состояния другого аллельного гена, то этодоминантный ген. При этом наблюдается полное проявление контролируемого доминантным геном признака, и сам признак называетсядоминантным. Примерами доминантных признаков служат: белый локон, брахидактилия, «куриная слепота», габсбургская губа, хондродистрофия; многочисленные формы доминантно наследуемых заболеваний. Если функциональное состояние одного аллельного гена зависит от состояния другого аллельного гена, то эторецессивныйген.Контролируемый этим геном
признак называетсярецессивным. Примерами рецессивных признаков служат: альбинизм, неспособность ощущать вкус фенилтиокарбамида (РТС):
многочисленные формы рецессивно наследуемых заболеваний.
В некоторых случаях доминантность и рецессивность относительны. Например, эпикант у монголоидов контролируется доминантным геном, а у бушменов и готтентотов — рецессивным геном.
Другой пример — ген плешивости, проявляющийся у мужчин как доминантный, а у женщин как рецессивный ген, что обусловлено действием гормонов. Это пример зависимого от пола наследования
—- кодоминирование: если аллельные гены в одинаковой мере активны, то это кодоминантные гены.
Например, известно, что серповидноклеточная анемия проявляется как аутосомно-рецессивное заболевание в гомозиготном организме. В таком организме присутствуют 2 патологических аллеля одно и того же гена, контролирующие синтез дефектного гемоглобина. В гетерозиготном организме присутствуют нормальный и дефектный гены. Причем ни один из этих генов не доминирует над другим. и в организме одновременно синтезируются оба вида гемоглобина (нормальный и дефектный). У таких индивидов симптоматики заболевания почти нет или она проявляется в очень легкой форме и лишь в условиях кислородной недостаточности. Этим лицам противопоказано проживание в высокогорных районах, служба в авиации, горноспасательная служба. Вместе с тем у них наблюдается высокая устойчивость к малярии (в 13 раз реже, чем в норме). Невосприимчивость к малярии у гетерозиготных носителей гена серповидноклеточной анемии объясняется неспособностью малярийного плазмодия осуществить свой жизненный цикл на дефектном гемоглобине.
|
|
Регуляция экспрессии гена.
Концепция оперона в регуляции экспрессии генов у прокариот. Ген обычно неактивен, но когда необходим определенный белок, конкретный ген «активируется», что обусловливает производство этого белка. Таким образом, клетки имеют механизм, контролирующий количество любого белка в определенное время. Синтез белков регулируется генетическим аппаратом, а также факторами внутренней и внешней среды.
Структура оперона прокариот. В 1961 г. два французских биолога Ф.Джакоб и Ж.Моно предложили механизм регуляции генов, названный гипотезой оперона.
Оперон - это последовательность специальных, функциональных сегментов ДНК, а также структурных генов, которые кодируют и регулируют синтез определенной группы белков одной метаболической цепи, например, ферментов гликолиза. Оперон (регулируемая единица транскрипции) состоит из следующих структурных частей (специальных последовательностей нуклеотидов) (слайд 2):
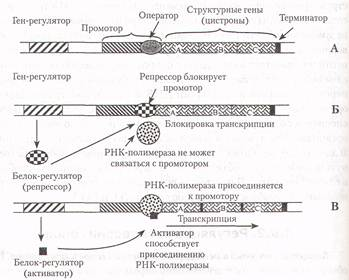
1. Ген-регулятор, контролирующий образование белка-регулятора.
2. Промотор - участок ДНК, к которому присоединяется РНК - полимераза и начинается транскрипция.
3. Оператор - участок промотора, связывающий белок-регулятор.
4. Структурные гены (цистроны) - участки ДНК, кодирующие мРНК конкретных белков.
5. Терминаторный участок ДНК несет сигнал об остановке транскрипции.