Цистеин
Цистеин способствует пищеварению, участвуя в процессах переаминирования. Способствует обезвреживанию некоторых токсических веществ и защищает организм от повреждающего действия радиации. Один из самых мощных антиоксидантов, при этом его антиоксидантное действие усиливается при одновременном приеме витамина С и селена. Цистеин является предшественником глутатиона – вещества, оказывающего защитное действие на клетки печени и головного мозга от повреждения алкоголем, некоторыми лекарственными препаратами и токсическими веществами, содержащимися в сигаретном дыме.
Содержание в продуктах питания
Сырая свинина, сырое куриное филе, сырое филе лосося, куриное яйцо, коровье молоко, семечки подсолнечника, грецкие орехи, мука пшеницы, кукурузная мука, неочищенный рис,соя сухая, горох и т.д.
Применение
Дополнительный прием цистеина необходим при ревматоидном артрите, заболеваниях артерий, раке. Он ускоряет выздоровление после операций, ожогов, связывает тяжелые металлы и растворимое железо. Эта аминокислота также ускоряет сжигание жиров и образование мышечной ткани.
При цистинурии, редком генетическом состоянии, приводящем к образованию цистиновых камней, принимать цистеин нельзя. Сахарный диабет также является противопоказанием для назначения цистеина.
При нарушении использования гомоцистеина в организме из него образуется гомоцистин. Гомоцистин может накапливаться в крови и тканях" выделяться с мочой, вызывая гомоцистинурию. Возможной причиной является наследственное нарушение обмена гомоцистеина либо гиповитаминоз фолиевой кислоты, а также витаминов В12 и В6. Патогенез Гомоцистеин, растворенный в плазме, провоцирует свободнорадикальное окисление липидов в липопротеинах крови и тем самым их задержку в крови, ускоряет агрегацию тромбоцитов, вызывает повреждение эндотелия сосудов. Сопутствующие заболевания Гомоцистеинемия считается фактором риска и обнаруживается в 30% случаев атеросклероза, тромбозов, ишемической болезни сердца. Она выявляется при болезни Альцгеймера, нарушениях беременности – невынашивание, мертворождения. Основы лечения При дефекте цистатионин-синтазы применяется лечение витамином В6 в дозе 250-500 мг/день. При дефекте метилен-тетрагидрофолат-редуктазы уровень гомоцистеина может быть снижен благодаря употреблению фолиевой кислоты по 5 мг/день. Витамин В12 также оказывает положительное влияние. Одновременно назначается диета со сниженным содержанием метионина, что достигается специальным подбором продуктов, бедных этой аминокислотой.
Из других биохимических нарушений можно отметить цистатионинурию, также часто возникающую при недостаточности витаминов группы В.
Биологические функции цистеина разнообразны и очень важны для организма. Так, цистеин, входящий в состав белков, играет необычайно важную роль в их фолдинге, поскольку тиогруппы цис способны образовывать прочную дисульфидную связь. При этом 2 остатка цистеина формируют молекулу цистина.
Окислительная реакция протекает либо с участием кофермента NAD+ под действием фермента цистеинредуктазы, либо неферментативно. Дисульфидные связи стабилизируют пространственную структуру полипептидной цепи или связывают между собой 2 цепи (например, А- и В-цепи гормона инсулина). Очень многие белки и ферменты в активном центре содержат SH-группы, участвующие в катализе. При их окислении ферментативная активность падает. Восстановление SH-групп часто происходит с использованием глутатиона - атипичного трипептида, содержащего γ-глутаминовую кислоту, цистеин и глицин.
Глутатион способен существовать в 2 формах - восстановленной (Г-SH) и окисленной (Г-S-S-F) и служит активным антиоксидантом в организме человека.
Как известно, некоторые ферменты содержат в активном центре SH-группы, абсолютно необходимые для каталитической реакции. При их окислении ферменты теряют свою активность. Предполагается, что одной из главных функций глутатиона является сохранение этих ферментов в активной восстановленной форме. Окисленный глутатион может вновь восстанавливаться под действием глутатионредуктазы, используя НАДФН2 и служить донатором водорода. С другой стороны, глутатион может оказывать ингибирующее действие на некоторые белки: в частности, известна реакция инактивации инсулина под действием глутатионинсулинтрансдегидрогеназы, в которой SH-глутатион является донатором водородных атомов, разрывающих дисульфидные связи между двумя полипептидными цепями молекулы инсулина. Показана коферментная функция глутатиона для дегидрогеназы фосфоглицериновой кислоты и глиоксалазы.
Ещё одним важным путём использования цис-теина можно считать синтез таурина в животных тканях, который происходит путём декарбоксилирования производных цистеина - цистеиновой и цистеинсульфиновой кислот.
Таурин необходим для синтеза парных жёлч ных кислот в печени. Кроме того, он очень важен в клетках как антиоксидант и используется для снижения ПОЛ и связывания гипохлоританиона (в форме хлораминового комплекса).
Цистеин также служит предшественником тиоэтаноламинового фрагмента HS-KoA (кофермента А).
Поскольку в процессе катаболизма сера метионина в тканях в основном переходит в серу цистеина и взаимопревращение цистина в цистеин легко осуществляется, проблема окисления серы всех аминокислот практически сводится к окислению цистеина. Главным путем оказался окислительный, включающий окисление цистеина в цистеинсульфиновую кислоту, трансаминирование последней с α-кетоглутаратом и образование пирувата и сульфита.
Сульфит, который получается в реакции, превращается в сульфат и выводится с мочой, либо превращается в эфиро-серные кислоты, которые также экскретируются почками. Цистеин - практически единственный источник сульфатов мочи.
Обмен серосодержащих аминокислот
В молекулах белка обнаружены три серосодержащие аминокислоты (ме-тионин, цистеин и цистин), метаболически тесно связанные друг с другом. Благодаря наличию в составе цистеина высокореактивной SH-группы в тканях легко осуществляется ферментативная окислительно-восстановительная реакция между цистеином и цистином.

Дисульфидная связь часто образуется между двумя остатками цистеина внутри одной полипептидной цепи или между двумя полипептидными цепями, способствуя тем самым стабилизации молекулы белка. Цистеинявляется составной частью трипептида глутатиона, сокращенно обозначаемого Г—SH, что подчеркивает функциональную значимость его тио-группы и возможность образования дисульфидной связи окисленного глу-татиона (Г—S—S—Г).
Известно, что многие ферменты содержат в активном центре SH-груп-пы, абсолютно необходимые для каталитической реакции. При их окислении ферменты теряют свою активность. Предполагают, что одной из главных функций глутатиона является сохранение этих ферментов в активной восстановленной форме. Окисленный глутатион может восстанавливаться под действием глутатионредуктазы, используя НАДФН. Кроме того, глутатион может оказывать ингибирующее действие на некоторые белки. В частности, известная реакция инактивации инсулина под действием глутатионинсулинтрансгидрогеназы, в которой SH-глутатион является донором водородных атомов, разрывающих дисульфидные связи между двумя полипептидными цепями молекулы инсулина. Установлена также коферментная функция глутатиона, в частности для глиоксилазы I. Ранее обсуждалось участие глутатиона в транспорте аминокислот через клеточную мембрану.
В процессе катаболизма сера метионина в тканях в основном переходит в серу цистеина, а взаимопревращение цистина в цистеин осуществляется легко. Поэтому проблема окисления серы всех аминокислот практически сводится к окислению цистеина. Главным путем оказался окислительный, включающий окисление цистеина в цистеинсульфиновую кислоту, транс-аминирование последней с α-кетоглутаратом и образование пирувата и сульфита по схеме:

Сульфит затем быстро окисляется в тканях и выводится с мочой в виде нетоксичных сульфатов и эфиросерных кислот. Использование цистеина и продуктов его окисления – цистеинсульфиновой и цистеиновой кислот – в образовании таурина рассмотрено ранее.
Метаболические пути превращения метионина в тканях значительно разнообразнее, чем пути превращения других серосодержащих аминокислот; тем не менее катаболизм метионина осуществляется через цистеин. Это превращение метионина в цистеин оказалось необратимым процессом. Выяснилось также, что углеродный скелет цистеина происходит из другой аминокислоты, а именно серина. Фактическим доноромметильных групп в реакциях трансметилирования является не свободный метионин, а так называемый активный метионин – S-аденозилметионин, который образуется в процессе АТФ-зависимой реакции, катализируемой метионин-аденозилтрансферазой.
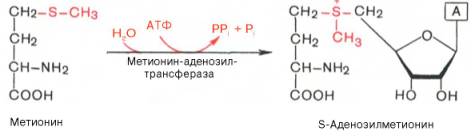
Своеобразие данной реакции заключается в том, что СН3-группа ме-тионина активируется под действием положительного заряда соседнего атома серы. S-аденозилметионин участвует во всех реакциях, где метильная группа используется в биосинтетических реакциях: например, в синтезе адреналина, креатинина, тимина, фосфатидилхолина, бетаина и др. Образовавшийся после отщепления метильной группы S-аденозилгомоцистеин подвергается гидролизу на аденозин и гомоцистеин; последний используется в синтезе серина (это основной путь превращения) или служит акцептором метильной группы от N5—СН3—ТГФК в синтезе метионина (эту реакцию катализирует гомоцистеинметилтрансфераза), завершая, таким образом, своеобразный цикл активирования метильной группы.

В качестве примера приводим схему биосинтеза креатина, в котором принимают участие три аминокислоты: аргинин, глицин и метионин. Реакция синтеза протекает в две стадии. Первая стадия – биосинтезгуанидинацетата – осуществляется в почках при участии глицин-амидинотранс-феразы (КФ 2.1.4.1):

Вторая стадия синтеза креатина протекает в печени при участии гуанидинацетатметилтрансферазы (КФ 2.1.1.2):
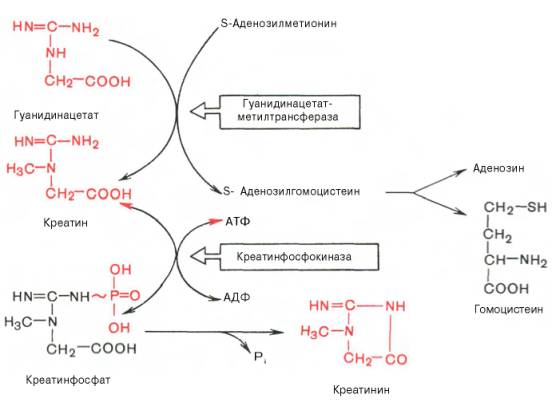
Креатин подвергается фосфорилированию с образованием креатин-фосфата, который последефосфорилирования (необратимая реакция) превращается в креатинин, выделяющийся с мочой.
Гомоцистеин может вновь превращаться в метионин путем метилирования. Однако основной путь дальнейшего превращения гомоцистеина связан с его использованием в синтезе цистеина, который может быть представлен в виде двух последовательных ферментативных реакций.

Ферменты, катализирующие синтез и распад цистатионина (циста-тионин-β-синтаза и цистатионаза), содержат ПФ. Цистеин далее подвергается окислению по описанному ранее пути, а гомосерин после транс-аминирования с α-кетоглутаратом превращается в α-кетомасляную кислоту; последняя может также образоваться из цистатионина непосредственно, минуя стадию гомосерина.