В ряде случаев, чтобы комплекс АГ-АТ стал крупнее, на эритроцитах адсорбируют АТ (антительный эритроцитарный диагностикум). В этом случае для исследования берут материал, в котором определяют наличие АГ. Как и в РНГА, в положительном случае образуется осадок в виде «зонтика», в отрицательном – в виде «пуговки».
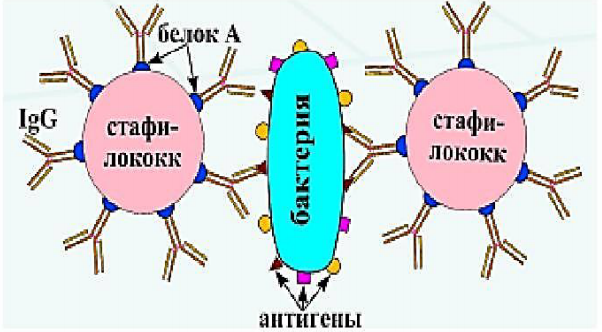
Реакция коагглютинации Применяют для определения АГ с помощьюантительного диагностикума – АТ,адсорбированных на белке А клеток стафилококка.Белок А стафилококка имеет сродство к Fc-фрагментуIg, поэтому такие бактерии, обработанные иммуннойдиагностической сывороткой, неспецифическиадсорбируют АТ сыворотки. Реакцию коагглютинацииставят на стекле. АТ диагностикума взаимодействуютантигенсвязывающими центрами с соответствующимимикроорганизмами, выделенными от больных. Врезультате коагглютинации образуются хлопья,состоящие из стафилококка, АТ диагностическойсыворотки и определяемого микроба.
Реакция Кумбса Реакция Кумбса – это разновидность реакцииагглютинации для определения антирезусных АТ.В сыворотке могут быть обнаруженыантирезусные АТ, которые являются неполными,одновалентными, т.е. способны связывать АГтолько одним активным центром. Ониспецифически взаимодействуют с АГ Rh+-эритроцитов, не вызывая их агглютинацию. Дляобнаружения таких АТ в систему «антирезусныеантитела + Rh+эритроциты» добавляютантиглобулиновую сыворотку (АТ противиммуноглобулинов человека), что вызываетагглютинацию эритроцитов.
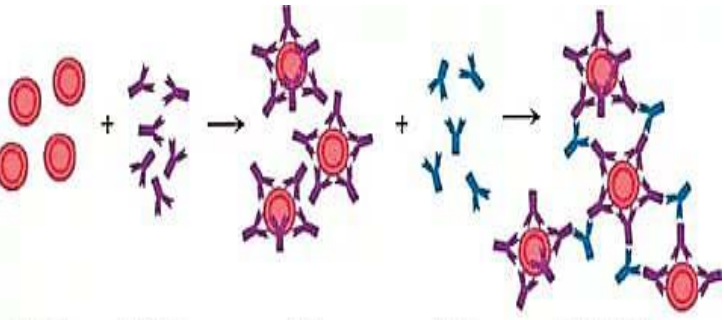
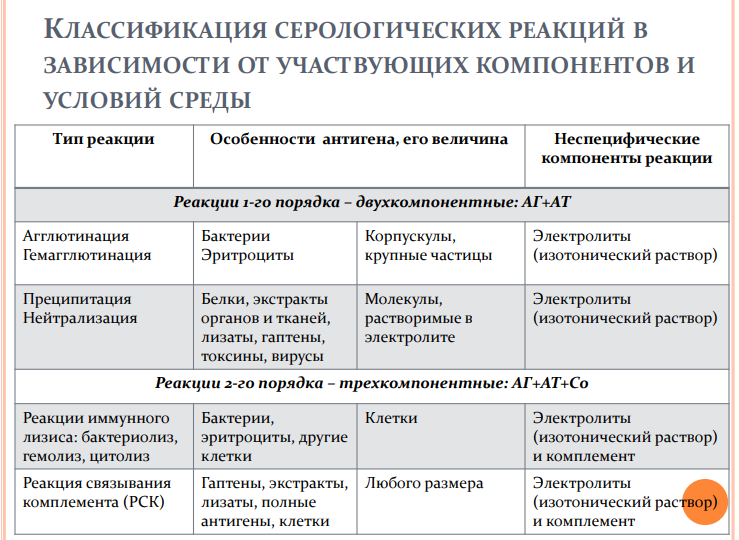
2. Реакции преципитации Реакция преципитации (от лат. praecipito –осаждать) – это формирование и осаждениекомплекса растворимого молекулярногоантигена с антителами в виде помутнения,называемого преципитатом. Механизмреакций преципитации также объясняетсятеорией «решетки». При этом АГ и АТ такжедолжны быть смешаны в эквивалентныхколичествах.
Реакция кольцепреципитации. Для этой реакции используют узкиепреципитационные пробирки диаметром 0,5см. В пробирку наливают диагностическую(т.е. содержащую антитела) сыворотку. Затемпо стенке пробирки на сыворотку медленнонаслаивают исследуемый антиген. Вположительном случае и при оптимальномсоотношении антигенов и антител на границерастворов образуется мутное кольцопреципитата, содержащее комплекс антигенантитело. Одновременно ставится контроль.
Реакция двойной иммунодиффузии по Оухтерлони. Разновидность реакции, в которой взаимодействиеантигена с антителом происходит не в жидкости, а вгеле. Чаще всего используют агаровый илиполиакриламидный гель.Для постановки реакции растопленный гельтонким слоем наносят на пластинку, послезатвердевания в нем вырезают лунки. Влунки отдельно помещают антигены ииммунные сыворотки, которыедиффундируют в гель. В положительномслучае на границе встречи образуетсяпреципитат в виде белой полосы. Умногокомпонентных систем между лунками сантигенами и антителами появляетсянесколько линий преципитата. У идентичныхантигенов линии преципитата сливаются, унеидентичных – пересекаются.
Реакция радиальной иммунодиффузии по Манчини. Иммунную сыворотку смешивают с расплавленнымагаровым гелем и равномерно наносят на стекло.После застывания в геле делают лунки, в которыепомещают антиген в различном разведении.Антиген, диффундируя в гель, образует сантителами кольцевые зоны преципитации вокруглунок. Диаметр кольца преципитациипропорционален концентрации антигена.Реакцию используют для определения в сывороткекрови иммуноглобулинов разных классов,компонентов системы комплемента и др.
Иммуноэлектрофорез. Представляет собой сочетание электрофореза вгеле с иммунодиффузией. Сначала проводятэлектрофоретическое разделение смесиантигенов в агаровом геле. Затем в канавку,параллельную направлению миграции белков,вносят диагностическую иммунную сыворотку.Антигены и антитела диффундируют в геле.В месте их взаимодействия образуютсядугообразные линии преципитации, количество,расположение и форма которых даютпредставление о составе исходной смесиантигенов.
Реакция преципитации для определения токсичности дифтерийной бактерии. Штаммы возбудителя дифтерии могут бытьтоксичными (продуцирующими экзотоксин) инетоксичными, что проверяется при диагностикезаболевания.Для этого в центр чашки с агаром кладут полоскуфильтровальной бумаги, пропитаннуюпротиводифтерийной антитоксической сывороткой.Параллельно полоске бляшками засеваютисследуемые штаммы бактерий и ставят в термостат(37о С, 24 ч.). В качестве контроля используют заведомотоксигенную культуру. Если бактерии выделяют токсин(является антигеном), то при его диффузии в гель наместе встречи с антителами сыворотки, которойпропитана бумага, образуется полоска преципитации.Если штамм нетоксигенный, то полоска не образуется.
II Реакции нейтрализации
1. Реакция нейтрализации in vivo. Проводится редко. Реакция направлена на выявлениеантител против токсина (антитоксина) у человека. Дляэтого в область предплечья внутрикожно вводятнебольшое количество токсина. Если покраснения иприпухлости в месте введения токсина нет, значит онбыл нейтрализован имеющимися антителами(положительная реакция, есть иммунитет). Еслинаблюдается местная реакция, значит токсиноказывает свое повреждающее действие,следовательно антител к нему нет (отрицательнаяреакция).Данная реакция была предложена Шиком длявыявления иммунитета к дифтерии.
2. Реакция нейтрализации in vitro. Реакции нейтрализации основаны на способности антигенов нейтрализовать in vitro биологически активные антигенсодержащие субстраты: токсины, вирусы, яды змей и т.п. Смешивают бав с сывороткой, содержащей антитела. Данную смесь вводят животным (в культуру клеток). Если животное остается здоровым, значит, произошла нейтрализация токсина антитоксином (антитоксическими антителами). Если у животного появляются симптомы интоксикации и оно погибает, то иммунный комплекс не образовался и не произошла нейтрализация
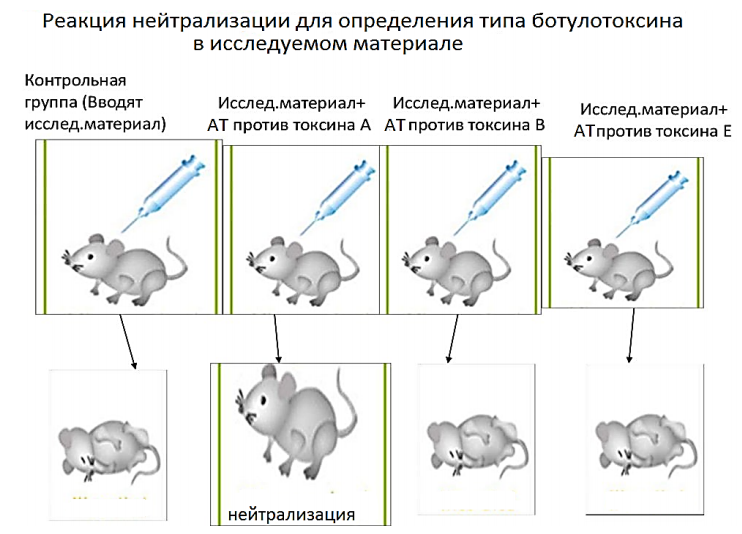
Пример, определение наличия ботулотоксина в материале. Исследуемый материал (сыворотка крови, фильтрат консервов и др.) вводят двум мышам: первой и второй (контрольная). При этом второй одновременно вводят сыворотку, содержащую антитела против всех типов данного токсина. Наблюдают. Если первая мышка погибла, а вторая осталась жива, значит в материале содержится ботулотоксин. Кроме наличия токсина, определяют и его тип (см. схему).
3. Реакция нейтрализации вирусов. Применяется для определения антител к вирусам. Смешивают сыворотку, исследуемую на наличие антител, с вирусом, к которому ищут антитела.
1) Данной смесью заражают чувствительных лабораторных животных. Через несколько суток регистрируют результаты. Если животное погибло, следовательно антител к данному вирусу в исследуемой сыворотке нет, нейтрализация не произошла. Если животное осталось живым и здоровым, следовательно произошла нейтрализация вируса имеющимися антителами (положительный результат).
2) Данной смесью заражают культуру клеток. При этом клетки находятся в питательной среде с индикатором на кислые продукты их метаболизма (изначально индикатор красного цвета, при наличии кислых метаболитов становится желтым). Если в сыворотке больного есть антитела, то они нейтрализуют вирус. При этом клетки остаются живыми и выделяют продукты жизнедеятельности, в результате индикатор меняет цвет на желтый (положительный результат). Если антител нет, то вирус приводит к гибели культуры клеток, в результате цвет среды остается красным. Реакцию также применяют для определения титра противовирусных антител: делают разведения сыворотки и выявляют, в каком титре антител достаточно для нейтрализации вируса.
4. Реакция торможения гемагглютинации (РТГА). Применяется для определения антител к вирусам, обладающим способностью склеивать эритроциты (Пр., вирус гриппа, кори, краснухи и др.). Делают последовательные разведения сыворотки больного в пробирках. Затем в каждую пробирку в одинаковом количестве добавляют взвесь эритроцитов и вирусы, к которым определяют антитела. Если антител достаточно для нейтрализации вируса, эритроциты остаются свободными и выпадают в осадок в виде «пуговки» (положительная реакция). Если сыворотка разведена настолько сильно, что антител в ней нет или недостаточно, вирус остается способным склеивать эритроциты, что приводит к образованию осадка в виде «зонтика».
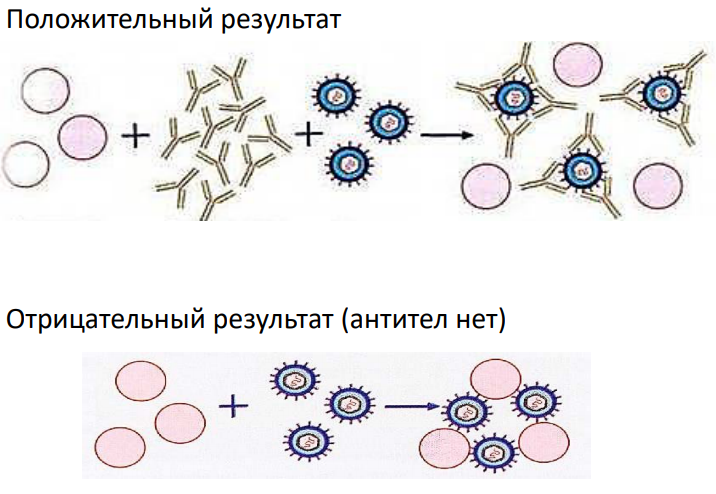
5. Реакция флоккуляции. Основана на способности токсина или анатоксина при смешивании в эквивалентных соотношениях с антитоксической сывороткой (содержит антитела против токсина) образовывать помутнение, а затем рыхлый осадок, похожий на комок шерсти – флоккулят. Механизм аналогичен реакции преципитации. Применяется для титрования антитоксических сывороток, токсинов и анатоксинов, определения типа токсина. *Анатоксин – это токсин, потерявший токсические свойства, но сохранивший антигенные.

Иммунологической памятью называют способность организма при повторной встрече с одним и тем же антигеном реагировать с более активным и более быстрым формированием иммунитета, т. е. реагировать по типу вторичного иммунного ответа. Повышенная чувствительность, или иммунореактивность, к антигену сохраняется при этом годами и даже десятилетиями. Иммунологическая память распространяется как на гуморальный (выработка антител), так и клеточный иммунитет. ГЗТ, трансплантационный иммунитет и другие формы иммунитета, связанные в основном с функцией лимфоцитов, «запоминаются» в не меньшей мере. Следовательно, иммунологическая память обусловлена деятельностью В-лимфоцитов (гуморальный иммунитет) и Т-лимфоцитов (клеточный иммунитет). Известно, что в популяции этих клеток имеются «клетки памяти», сохраняющие многие годы способность реагировать на повторное введение антигена, так как вырабатывают рецепторы к этому антигену. Видимо, ведущая роль в сохранении иммунологической памяти используется в практике вакцинации людей. Это делают в тех случаях, когда необходимо создать высокий уровень иммунитета и поддерживать его длительное время на этом уровне. Осуществляют это 2-3-кратными прививками при первичной вакцинации и периодическими повторными прививками – ревакцинациями. Например, против дифтерии прививают ребенка на втором году жизни, затем в детском саду, школе, армии, т. е. прививки проводят с интервалами в 5-7 лет. Ревакцинацию применяют также по противоэпидемическим показаниям, когда необходимо быстро создать коллективный иммунитет среди ранее прививавшихся людей (например, при дифтерии, брюшном тифе, чуме и других инфекциях).
Первичный иммунный ответ. Индуктивная фаза – до 2 суток. Первые антитела начинают обнаруживаться в сыворотке с 3-го дня после контакта с антигеном. Первые антитела – IgМ C 5-7 дня – постепенная смена на IgG той же специфичности. К 12-15 дню концентрация антител в сыворотке достигает максимума, стационарная фаза продолжается 15-30 суток, а фаза снижения – 1-6 мес. Но некоторое кол-во антител можно обнаружить через несколько месяцев и даже лет, благодаря ИММУНОЛОГИЧЕСКОЙ ПАМЯТИ.
По окончании ИО наряду с эффекторными клетками формируются Т- и В- клетки памяти. Это долгоживущие ЛФ (срок жизни – 10 и более лет). Существуют CD4+ и CD8+ Тклетки памяти, В-клетки памяти и долгоживущие плазматические клетки.
Антитела к некоторым антигенам определяются в сыворотке на протяжении нескольких десятков лет, хотя период полураспада самых стойких иммуноглобулинов составляет 25 дней. В организме происходит постоянная выработка антител заданной специфичности и сохранение их в качестве рецепторов на клетках памяти (преимущественно IgG и IgA
Вторичный иммунный ответ. Развивается быстрее. Требует меньших доз антигена. Проявления более интенсивны. Специфичность гуморальных и клеточных реакций выше. Обеспечивает более эффективную защиту.
Вторичный иммунный ответ. При повторном контакте с антигеном происходит его комплементарное соединение с рецептором на клетках памяти. Происходит селекция лимфоцитов, имеющих на мембране соответствующие рецепторы, в результате образуется клон определенных лимфоцитов, несущих на мембране комплекс «АГ+ГКГС 2 класса+рецептор (Ig)», который распознается всеми иммунокомпетентными клетками.
Индуктивная фаза составляет несколько часов. Продуктивная фаза протекает быстрее и интенсивнее – антитела в сыворотке обнаруживаются уже с первых суток. Вырабатываются преимущественно IgG. Пиковая концентрация антител достигается уже к 3-4 суткам и долго не снижается.
Переключение синтеза классов антител находится под контролем Т-клеток. Для осуществления переключения необходимы межклеточный контакт, опосредованный взаимодействием CD40 В- и Т-клеток, а также действием цитокинов:
ИЛ-4 для переключения на синтез IgG1 и IgE;
γ-ИФН для переключения на синтез IgG2а и IgG3;
Трансформирующий фактор роста β для переключения на синтез IgG2b и IgА;
ИЛ-6 усиливает продукцию любых антител;
ИЛ-5 стимулирует синтез IgА;
ИЛ-4 усиливает продукцию IgE
Гуморальный ИО на Т-независимые антигены заключается в образовании в основном IgМ, переключение синтеза может отсутствовать; При ИО на Т-зависимые антигены происходит переключение синтеза АТ под влиянием ИЛ-1 и ИЛ-3, ГМ-КСФ), контакта с макрофагами и некоторыми другими клетками.
Вторичный Т-клеточный ответ также протекает быстрее и интенсивнее, чем первичный; Для Т-клеток памяти достачно распознавание комплекса антигенного пептида с молекулой ГКГС 1 класса в присутствии ИЛ-2; Дополнительной костимуляции не требуется

Анамнестическая реакция, реакция, возникающая в иммунизированном или перенесшем инфекционную болезнь организме, утратившем способность сохранять антитела. На анамнестической реакции основано применение повторных прививок.
Титр антител — предельное разведение сыворотки крови, при котором могут быть обнаружены антитела. Позволяет оценить количество и разнообразие антител в крови человека и коррелирующую с ними силу иммунного ответа организма. Для измерения титра антител используется иммуноферментный анализ (ИФА) — наиболее доступный метод диагностики.
Иммуноглобулины по структуре, антигенным и иммунобиологическим свойствам разделяются на пёять классов: IgM, IgG, IgA, IgE, IgD.
Иммуноглобулин класса G. Изотип G составляет основную массу Ig сыворотки крови. На его долю приходится 70—80 % всех сывороточных Ig, при этом 50 % содержится в тканевой жидкости. Среднее содержание IgG в сыворотке крови здорового взрослого человека 12 г/л. Период полураспада IgG — 21 день.
IgG — мономер, имеет 2 антигенсвязывающих центра (может одновременно связать 2 молекулы антигена, следовательно, его валентность равна 2), молекулярную массу около 160 кДа и константу седиментации 7S. Различают подтипы G1, G2, G3 и G4. Синтезируется зрелыми В-лимфоцитами и плазматическими клетками. Хорошо определяется в сыворотке крови на пике первичного и при вторичном иммунном ответе.
Обладает высокой аффинностью. IgG1 и IgG3 связывают комплемент, причем G3 активнее, чем G1. IgG4, подобно IgE, обладает цитофильностью (тропностью, или сродством, к тучным клеткам и базофилам) и участвует в развитии аллергической реакции I типа. В иммунодиагностических реакциях IgG может проявлять себя как неполное антитело.
Легко проходит через плацентарный барьер и обеспечивает гуморальный иммунитет новорожденного в первые 3—4 месяца жизни. Способен также выделяться в секрет слизистых, в том числе в молоко путем диффузии.
IgG обеспечивает нейтрализацию, опсонизацию и маркирование антигена, осуществляет запуск комплемент-опосредованного цитолиза и антителозависимой клеточно-опосредованной цитотоксичности.
Иммуноглобулин класса М. Наиболее крупная молекула из всех Ig. Это пентамер, который имеет 10 антигенсвязывающих центров, т. е. его валентность равна 10. Молекулярная масса его около 900 кДа, константа седиментации 19S. Различают подтипы M1 и М2. Тяжелые цепи молекулы IgM в отличие от других изотипов построены из 5 доменов. Период полураспада IgM — 5 дней.
На его долю приходится около 5—10 % всех сывороточных Ig. Среднее содержание IgM в сыворотке крови здорового взрослого человека составляет около 1 г/л. Этот уровень у человека достигается уже к 2—4-летнему возрасту.
IgM филогенетически — наиболее древний иммуноглобулин. Синтезируется предшественниками и зрелыми В-лимфоцитами. Образуется в начале первичного иммунного ответа, также первым начинает синтезироваться в организме новорожденного — определяется уже на 20-й неделе внутриутробного развития.
Обладает высокой авидностью, наиболее эффективный активатор комплемента по классическому пути. Участвует в формировании сывороточного и секреторного гуморального иммунитета. Являясь полимерной молекулой, содержащей J-цепь, может образовывать секреторную форму и выделяться в секрет слизистых, в том числе в молоко. Большая часть нормальных антител и изоагглютининов относится к IgM.
Не проходит через плаценту. Обнаружение специфических антител изотипа М в сыворотке крови новорожденного указывает на бывшую внутриутробную инфекцию или дефект плаценты.
IgM обеспечивает нейтрализацию, опсонизацию и маркирование антигена, осуществляет запуск комплемент-опосредованного цитолиза и антителозависимой клеточно-опосредованной цитотоксичности.
Иммуноглобулин класса А. Существует в сывороточной и секреторной формах. Около 60 % всех IgA содержится в секретах слизистых.
Сывороточный IgA: На его долю приходится около 10—15% всех сывороточных Ig. В сыворотке крови здорового взрослого человека содержится около 2,5 г/л IgA, максимум достигается к 10-летнему возрасту. Период полураспада IgA — 6 дней.
IgA — мономер, имеет 2 антигенсвязывающих центра (т. е. 2-валентный), молекулярную массу около 170 кДа и константу седиментации 7S. Различают подтипы А1 и А2. Синтезируется зрелыми В-лимфоцитами и плазматическими клетками. Хорошо определяется в сыворотке крови на пике первичного и при вторичном иммунном ответе.
Обладает высокой аффинностью. Может быть неполным антителом. Не связывает комплемент. Не проходит через плацентарный барьер.
IgA обеспечивает нейтрализацию, опсонизацию и маркирование антигена, осуществляет запуск антителозависимой клеточно-опосредованной цитотоксичности.
Секреторный IgA: В отличие от сывороточного, секреторный sIgA существует в полимерной форме в виде ди- или тримера (4- или 6-валентный) и содержит J- и S-пeптиды. Молекулярная масса 350 кДа и выше, константа седиментации 13S и выше.
Синтезируется зрелыми В-лимфоцитами и их потомками — плазматическими клетками соответствующей специализации только в пределах слизистых и выделяется в их секреты. Объем продукции может достигать 5 г в сутки. Пул slgA считается самым многочисленным в организме — его количество превышает суммарное содержание IgM и IgG. В сыворотке крови не обнаруживается.
Секреторная форма IgA — основной фактор специфического гуморального местного иммунитета слизистых оболочек желудочно-кишечного тракта, мочеполовой системы и респираторного тракта. Благодаря S-цепи он устойчив к действию протеаз. slgA не активирует комплемент, но эффективно связывается с антигенами и нейтрализует их. Он препятствует адгезии микробов на эпителиальных клетках и генерализации инфекции в пределах слизистых.
Иммуноглобулин класса Е. Называют также реагином. Содержание в сыворотке крови крайне невысоко — примерно 0,00025 г/л. Обнаружение требует применения специальных высокочувствительных методов диагностики. Молекулярная масса — около 190 кДа, константа седиментации — примерно 8S, мономер. На его долю приходится около 0,002 % всех циркулирующих Ig. Этот уровень достигается к 10—15 годам жизни.
Синтезируется зрелыми В-лимфоцитами и плазматическими клетками преимущественно в лимфоидной ткани бронхолегочного дерева и ЖКТ.
Не связывает комплемент. Не проходит через плацентарный барьер. Обладает выраженной цитофильностью — тропностью к тучным клеткам и базофилам. Участвует в развитии гиперчувствительности немедленного типа — реакция I типа.
Иммуноглобулин класса D. Сведений об Ig данного изотипа не так много. Практически полностью содержится в сыворотке крови в концентрации около 0,03 г/л (около 0,2 % от общего числа циркулирующих Ig). IgD имеет молекулярную массу 160 кДа и константу седиментации 7S, мономер.
Не связывает комплемент. Не проходит через плацентарный барьер. Является рецептором предшественников В-лимфоцитов.
3. Аллергический метод диагностики инфекционных заболеваний. Фазы, механизм, защитная и патогенетическая роль инфекционной аллергии (ГЧЗТ). Диагностические препараты для постановки кожно-аллергических проб (корпускулярные и растворимые.
I тип - анафилактический.
При первичном контакте с антигеном образуются IgE, которые прикрепляются Fc-фрагментом к тучным клеткам и базофилам. Повторно введенный антиген перекрестно связывается с IgE на клетках, вызывая их дегрануляцию, выброс гистамина и других медиаторов аллергии.
II тип - цитотоксический.
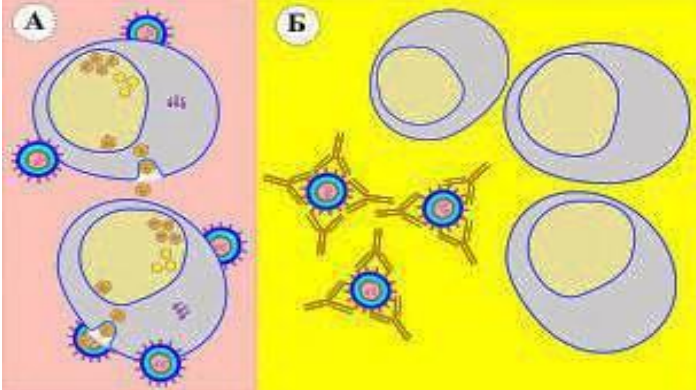
Антиген, расположенный на клетке "узнается" антителами классов IgG, IgM. При взаимодействии типа "клетка-антиген-антитело", происходит активация комплемента и разрушение клетки по трем направлениям: комплемент-зависимый цитолиз (А); Фагоцитоз (Б); антителозависимая клеточная цитотоксичность (В).
III тип - иммунокомплексный.
Антитела классов IgG, IgM образуют с растворимыми антигенами иммунные комплексы, которые активируют комплемент. При избытке антигенов или недостатке комплемента иммунные комплексы откладываются на стенке сосудов, базальных мембранах, т.е. структурах, имеющих Fс-рецепторы.
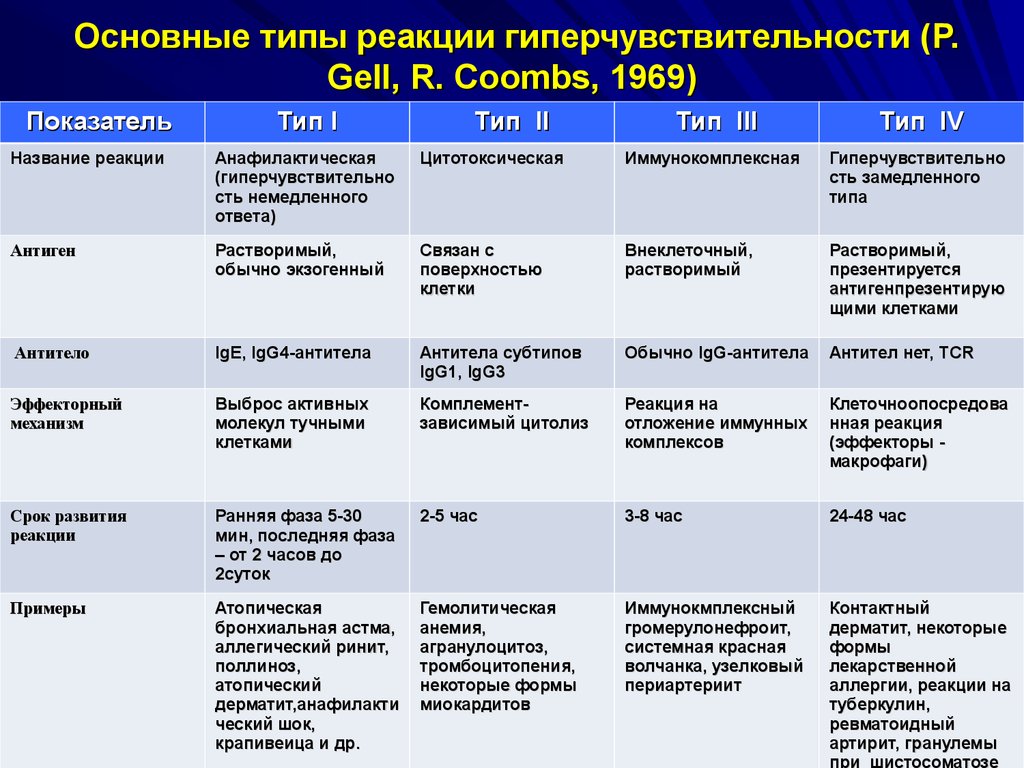
IV тип - гиперчувствительность замедленного типа.
Этот тип обусловлен взаимодействием антигена с макрофагами и Thl-лимфоцитами, стимулирущими клеточный иммунитет (клеточно-опосредованная или туберкулиновая гиперчувствительность.
ГЗТ обусловлена макрофагами, Тн1-лимфоцитами и ЦТЛ.
• Тн1-хелперы и ЦТЛ секретируют интерферон γ ®активация макрофагов®индукция воспаления.
• Макрофаги вовлекаются в процесс разрушения антигена.
• После первичного попадания аллергена в организме образуется клон Т-лимфоцитов, несущих рецепторы, распознающие этот аллерген.
• При повторном попадании Т-лимфоциты взаимодействуют с аллергеном, активируются и выделяют цитокины, которые вызывают хемотаксис макрофагов и активируют их.
• Активированные макрофаги выделяют множество веществ, которые вызывают воспаление и уничтожают аллерген.
• Возникает через 1 – 3 суток после повторного воздействия аллергена, т.к. для развития реакции необходимо определенное количество Т-клеток, на что требуется 24-72 часа.
• Морфологическая картина ГЗТ: уплотнение и воспаление ткани в результате её инфильтрации Т-лимфоцитами и макрофагами.
В основе этого типа гиперчувствительности лежит взаимодействие цитотоксического (сенсибилизированного) Т-лимфоцита со специфическим антигеном, что приводит к высвобождению из Т-клетки целого набора цито-кинов, опосредующих проявления замедленной гиперчувствительности.
Клеточный механизм включается при:
1. Недостаточной эффективности гуморального механизма (например, при внутриклеточном расположении возбудителя – туберкулезной палочки, бруцеллы);
2. В том случае, когда в роли антигена выступают чужеродные клетки (некоторые бактерии, простейшие, грибы, пересаженные клетки и органы), либо клетки собственных тканей, антигены которых изменены (например, включение аллергена-гаптена в белки кожи и развитие контактного дерматита).
Таким образом, в период иммунологической стадии в организме созревают цитотоксические (сенсибилизированные) Т-лимфоциты.
Во время повторного контакта с антигеном (аллергеном), в патохимической стадии цитоток-сические (сенсибилизированные) Т-лимфоциты выделяют следующие цитокины:
1. Фактор, угнетающий миграцию макрофагов (МИФ, MIF), который обладает способностью усиливать фагоцитоз и участвует в образовании гранулем;
2. Фактор, стимулирующий образование эндогенных пирогенов (ИЛ-1);
3. Митогенные (ростовые) факторы (ИЛ-2, ИЛ-3, ИЛ-6 и т.д.);
4. Хемотаксические факторы для каждой линии белых клеток, особенно, ИЛ-8;
5. Гранулоцитарно-моноцитарные колониестимулирующие факторы;
6. Лимфотоксины;
7. Опухоль-некротизирующий фактор;
8. Интерфероны (альфа, бета, гамма).
Выделяющиеся из сенсибилизированных Т-лимфоцитов цитокины активируют и привлекают в очаг воспаления клетки моноцитарно-макрофагального ряда.
В том случае, если действие лимфоцитов направлено против вирусов, инфицирующих клетки, или против трансплантационных антигенов, стимулированные Т-лимфоциты трансформируются в клетки, обладающие свойствами клеток-киллеров по отношению к клеткам-мишеням, несущим данный антиген. К таким реакциям относятся: аллергия, формирующаяся при некоторых инфекционных заболеваниях, реакция отторжения трансплантата, некоторые виды аутоиммунных поражений. На рис. 57 представлена схема аллергической реакции IV (замедленного) типа.
Таким образом, во время патофизиологической стадии повреждение клеток и тканей происходит за счет:
· Прямого цитотоксического действия Т-лимфоцитов;
· Цитотоксического действия Т-лимфоцитов за счет неспецифических факторов (провос-палительные цитокины, апоптоз и др.);
· Лизосомальных ферментов и иных цитотоксических веществ (NO, оксиданты) активированных клеток моноцитарно-макрофагального ряда.
При аллергических реакциях IV типа среди клеток, инфильтрирующих очаг воспаления, преобладают макрофаги, затем Т-лимфоциты и, в последнюю очередь, нейтрофилы.
Примером гиперчувствительности замедленного типа являются аллергический контактный дерматит, реакция отторжения аллотрансплантата, туберкулез, лепра, бруцеллез, микозы, про-тозойные инфекции, некоторые аутоиммунные заболевания.
Формы ГЗТ
• -туберкулиновая реакция;
• -контактная аллергия;
• - гранулематозная аллергия (замедленная аллергия к белкам).
Туберкулиновая проба - Покраснение, припухлость и уплотнение на месте введения аллергена. Возникает через 5-6 час после внутрикожного введения туберкулина и достигает максимума через 48-72 часа. Реакция специфична!!!