Биполярные, горизонтальные и амакриновые клетки
Общая схема связей между клетками сетчатки была составлена на основе морфологических описаний Рамон-и-Кахаля: проводящий путь от фоторецепторов до ганглиозных клеток при помощи биполярных клеток, с регулированием передачи посредством горизонтальных и амакриновых клеток. Исходя из схемы соединений клеток в сетчатке приматов становится понятным, что конечный выходной сигнал является результатом сложного интегративного процесса, происходящего в самой сетчатке. Например, горизонтальная клетка, показанная на рис.2D, получает синаптические входы от многих фоторецепторов и, в свою очередь, образует с ними модулирующую обратную связь.

Рис. 1. Основные клеточные типы и соединения в сетчатке приматов представлены здесь для иллюстрации связей между фоторецепторами (палочками и колбочками) и ганглиозными клетками.
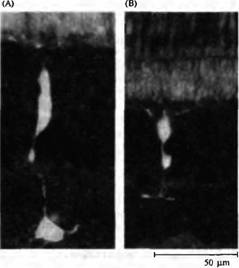

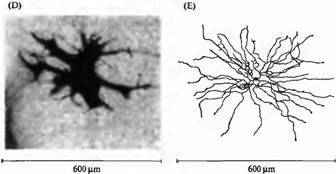
Рис. 2. Биполярные, горизонтальные и амакри новые клетки. (А) Деполяризующаяся биполярная клетка "оп"-центра золотой рыбки с введенным флуоресцентным красителем. (В) Гиперполяризующаяся биполярная клетка "off''-центра золотой рыбки. (С) Выделенная биполярная клетка из сетчатки крысы, окрашенная на про теинкиназу С. (D) Горизонтальная клетка акулы (морской собаки) с введенной пероксидазой хрена. (Е) Накапливающая индоламин амакриновая клетка из сетчатки кролика с введенным красителем Lucifer Yellow.
Горизонтальные клетки также образуют окончания на биполярных клетках. Аналогично некоторые амакриновые клетки (рис.2Е), получающие сигналы от биполярньи клеток, посылают аксоны как обратно на биполярные, так и на ганглиозные клетки. Можно сделать вывод, что горизонтальные и амакриновые клетки осуществляют и модулируют процесс передачи сигнала в сетчатке. Дополнительную сложность создает то, что каждый из основных классов нейронов, показанных на рис.1 и 2, имеет множество морфологических и фармакологических подтипов). При помощи электрофизиологических, биохимических и анатомических критериев было описано несколько основных классов биполярньи клеток, больше чем 2 типа горизонтальных клеток и по крайней мере 20 типов амакриновых клеток.
Медиаторы в сетчатке

Рис. 3. Лентовидные синапсы, образованные терминалью фоторецептора на окончаниях биполярных и горизонтальных клеток. Пресинаптические везикулы в терминалях колбочек (С) выстроены вдоль ленты (R). Данный тип синапсов приспособлен к постоянному высвобождению квантов глутамата к биполярным (В) и горизонтальным (Н) клеткам в темноте. Через ГАМК-ергические горизонтальные клетки осуществляется обратная связь с фоторецепторами.
В сетчатке были обнаружены практически все известные медиаторы. Глутамат высвобождается фоторецепторами и биполярными клетками. Горизонтальные клетки секретируют ГАМК. Некоторые амакриновые клетки секретируют дофамин, другие ацетилхолин или индоламины. Распределение и функциональная значимость этих медиаторов, рецепторов и их субъединиц, а также переносчиков сигнала были детально изучены методами иммуногистохимии и гибридизации in situ. Некоторые медиаторы-пептиды, например VIP (вазоактивный интестинальный пептид), обнаруженные в глазу, могут играть важную трофическую роль в развитии зрительной системы и быть связаны с развитием миопии (близорукости).
Постоянное высвобождение квантов глутамата фоторецепторами в темноте осуществляется при помощи специального типа нервных окончаний, называемых лентовидными синапсами (ribbon synapses). Пример такого синапса показан на рис.3. Терминаль колбочки содержит везикулы, плотно связанные с мембраной и готовые к высвобождению, которые расположены вдоль длинной плоской органеллы, называемой "лентой" (ribbon). Постсинаптическая мембрана образована терминалями биполярных и горизонтальных клеток, которые формируют инвагинации на окончаниях фоторецепторов.
Концепция рецептивных полей
Техника освещения определенных зон сетчатки привела к появлению такой важной концепции, как рецептивное поле, которая стала ключом к пониманию значения сигналов не только на уровне сетчатки, но и на последующих стадиях в коре. Сам термин "рецептивное поле" был исходно введен Шеррингтоном по отношению к рефлексам и был перенесен в зрительную систему Хартлайном.
Рецептивное поле нейрона зрительной системы может быть определено как зона сетчатки, при падении света на которую может изменяться активность данного нейрона.
Например, регистрация активности одного нейрона зрительного нерва и коры кошки показала, что частота его импульсации увеличивается или уменьшается, если меняется освещение над определенной зоной сетчатки.
Эта зона является рецептивным полем данного нейрона.
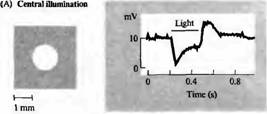
---------

Рис. 4. Карта рецептивных полей биполярных клеток, чувствительных к гиперполяризации (Н). Сигналы регистрировались на биполярной клетке сетчатки золотой рыбки, которая отвечала гиперполяризацией (А) на освещение центра рецептивного поля. Освещение в виде кольца вызывало деполяризацию клетки (В). Рассеянный свет слабо влиял на потенциал клетки. Для биполярной клетки, чувствительной к деполяризации (D), освещение центра рецептивного поля приводит к деполяризации, в то время как освещение в виде кольца вызывает гиперполяризацию.
По определению освещение за пределами рецептивного поля не оказывает влияния на импульсацию. Эта зона сама по себе может быть подразделена на несколько различных регионов, освещение некоторых из которых увеличивает активность, а других подавляет. Это описание рецептивных полей также применимо по отношению к другим нейронам, таким как биполярные и горизонтальные клетки, у которых локальный ответ переменной амплитуды вызывается падением света на сетчатку (что обсуждается далее в этой главе). Как будет показано далее, отдельные вспышки ненаправленного света практически бесполезны при оценке функции зрительной системы.
Ответы биполярных клеток
Каждая биполярная клетка получает сигналы непосредственно от палочки либо от колбочки. Клетки обычно получают сигналы от 15-45 рецепторов. Существует особый тип биполярных клеток, так называемые "карликовые биполяры" (midget bipolar), которые получают сигнал только от одной колбочки. Логично ожидать, что большинство карликовых биполяров находится в области ямки, где острота зрения максимальна. Они передают сигналы с фоторецептора на специальный тип ганглиозных клеток. В других отделах сетчатки биполярные клетки собирают информацию с 15-20 колбочек. Биполярные и горизонтальные клетки отвечают на освещение градуальной де - или гиперполяризацией.
Сигналы и рецептивные поля биполярных клеток зависят от двух механизмов. Во--первых, постоянное высвобождение медиатора в темноте способствует поддержанию ряда биполярных клеток в состоянии постоянной деполяризации, а других - в состоянии гиперполяризации, в зависимости от того, какие - возбуждающие или тормозные - рецепторы они имеют. Во-вторых, свет приводит к гиперполяризации фоторецепторов, тем самым уменьшая выброс глутамата. Уменьшение постоянного освобождения глутамата фоторецепторами при освещении приводит, соответственно, к уменьшению активности биполярных клеток, имеющих возбуждающие рецепторы глутамата, т.е. вызывают их гиперполяризацию. Такие клетки называются Н-биполяры (hyperpolarizing).
Наоборот, уменьшение тонического высвобождения глутамата фоторецепторами при освещении последних приводит к деполяризации тех биполярных клеток, которые экспрессируют тормозные рецепторы глутамата - это D-биполяры (depolarizing). D-биполярные клетки являются одним из редких типов клеток, у которых были обнаружены тормозные эффекты, опосредуемые через рецепторы глутамата. Канеко с коллегами показали, что данный тормозный эффект обусловлен метаботропными рецепторами глутамата, расположенными на мембранах биполярных клеток, которые действуют через G-белок и вторичные посредники, приводя к уменьшению ионной проводимости. Таким образом, в темноте в результате активации этих рецепторов ионные каналы, управляемые циклическими нуклеотидами на D-биполярных клетках, закрываются (в отличие от каналов фоторецепторов). После освещения высвобождение глутамата уменьшается, открываются катионные каналы, и биполярные клетки деполяризуются.