Снаружи каждая клетка одета нежным эластичным покровом, который составляет неотъемлемый функциональный компонент клетки и называется плазматической мембраной. Эта мембрана играет чрезвычайно важную роль в регулировании состава клеточного содержимого, так как через нее в клетку поступают все питательные вещества и выходят наружу все отходы или продукты секреции. Мембрана задерживает проникновение в клетку одних веществ и облегчает поступление других. Клетки почти всегда окружены водной средой; это может быть пресная или морская вода (в случае простейших организмов), тканевый сок (высшие растения), плазма или внеклеточная жидкость (высшие животные).
Плазматическая мембрана действует так, как будто она пронизана ультрамикроскопическими порами, через которые проходят определенные вещества, причем величиной этих пор определяется максимальная величина способных пройти через них молекул. Возможность прохождения вещества через мембрану зависит не только от величины молекул, но и от электрического заряда диффундирующей частицы (если она им обладает), от присутствия и числа молекул воды, связанных с поверхностью этих частиц, и от растворимости частиц в липидах. Химическая и физическая природа мембраны выяснена еще не до конца, но, по-видимому, это трехслойная пленка толщиной около 12 нм. Наружный и внутренний слои, каждый толщиной около 3 нм, состоят из белка, а между ними лежит слой фосфолипидных молекул, имеющий толщину 60 нм.
Такого рода трехслойную структуру можно видеть на электронных микрофотографиях, полученных при высоком разрешении. Интересно, что все плазматические мембраны животных, растительных и бактериальных клеток, так же как и мембраны разнообразных внутриклеточных органелл, имеют, видимо, сходное трехслойное строение. Два слоя белка, разделенные слоем липида, так называемая элементарная мембрана, по-видимому, представляет собой широко распространенную основную единицу мембранной структуры.
У растений почти все клетки имеют, кроме того, толстую клеточную стенку, состоящую из целлюлозы и лежащую кнаружи от плазматической мембраны (у большинства животных клеток ее нет). Клеточная стенка во многих местах пронизана мельчайшими отверстиями, через которые протоплазма одной клетки соединяется с протоплазмой других, соседних с ней клеток; через эти отверстия вещества могут переходить из одной клетки в другую. Плотные, прочные клеточные стенки создают опору телу растения.
Для того чтобы понять механизмы, лежащие в основе обмена материалами между клеткой и окружающей средой, мы должны прежде всего учитывать, что для всех молекул в жидкостях и газах характерна тенденция диффундировать, т. е. перемещаться во всех направлениях до тех пор, пока они не распределятся равномерно по всему доступному пространству. Диффузию можно определить как распространение молекул из области их высокой концентрации в область более низкой концентрации, обусловленное их тепловым движением (рис. 19). Скорость диффузии зависит от величины молекул и от температуры. Молекулы, из которых построены все вещества, в том числе и твердые, находятся в постоянном движении. Основное различие между тремя состояниями вещества — твердым, жидким и газообразным — определяется степенью свободы движения молекул. Молекулы твердого тела упакованы сравнительно плотно, и силы притяжения между ними позволяют им совершать колебания, но не допускают свободного передвижения. В жидкости расстояния между молекулами несколько больше, межмолекулярные силы слабее и молекулы обладают значительной свободой передвижения. Наконец, в газообразном веществе молекулы настолько удалены друг от друга, что межмолекулярные силы ничтожны и свободу движения молекул ограничивают только внешние препятствия.
Если исследовать каплю воды под микроскопом, то движение молекул обнаружить не удастся, но если добавить к ней капельку туши (которая содержит мелкие частицы угля), то можно наблюдать непрерывное беспорядочное движение частиц угля. Каждая частица угля непрерывно подвергается ударам молекул воды, и толчки от этих ударов приводят эти частицы в движение. Такое движение мелких частиц называют броуновским движением по имени Роберта
Броуна — английского ботаника, который его впервые заметил, рассматривая под микроскопом пыльцевые зерна в капле воды. В процессе диффузии каждая молекула движется по прямой линии, пока не столкнется с чем-нибудь, например с другой молекулой или со стенкой сосуда; тогда она отскакивает и начинает перемещаться в другом направлении. Молекулы продолжают двигаться и после того, как они равномерно распределились по всему доступному пространству; однако, в то время как одни молекулы переходят, например, слева направо, другие переходят справа налево, и равновесие сохраняется. Различные вещества (сколько бы их ни было), находящиеся в одном и том же растворе, диффундируют независимо друг от друга. Скорость движения отдельных молекул может достигать нескольких сот метров в секунду, но каждая молекула до столкновения с другой, от которой она отскакивает, проходит лишь малую долю нанометра. Поэтому перемещение молекулы в каком-нибудь одном направлении происходит очень медленно. В этом можно убедиться, положив на дно стеклянного цилиндра, наполненного водой, кусочек какой-нибудь краски. Через несколько дней можно заметить, что красящее вещество постепенно поднимается вверх, но пройдут месяцы, прежде чем краска равномерно распределится по всему цилиндру. Таким образом, хотя диффузия на очень малые расстояния происходит весьма быстро, для того чтобы пройти расстояние в несколько сантиметров, молекулам требуется много времени.
Этот факт имеет большое биологическое значение, так как из него следует, что число молекул кислорода или питательных веществ, которые могут достигнуть организма путем одной только диффузии, весьма ограничено. Лишь очень небольшой организм, которому ежесекундно необходимо относительно небольшое число молекул питательных веществ или кислорода, может выжить, сидя на одном месте и дожидаясь, когда эти молекулы дойдут до него путем диффузии. Более крупный организм должен иметь возможность либо передвигаться с места на место, либо приводить в движение окружающую среду и таким путем доставлять себе нужные молекулы, либо, наконец, он может обитать в таком месте, где сама среда непрерывно движется, например в реке или на морском побережье в зоне приливов. Крупные наземные растения — деревья и кустарники — разрешили эту проблему особым образом: у них имеется чрезвычайно сильно разветвленная корневая система, при помощи которой они получают необходимые им вещества с большого участка окружающей среды.
Могут ли молекулы данного вещества проходить через ту или иную мембрану, зависит от ее структуры и от величины имеющихся в ней пор. Мембрану называют проницаемой, если через нее проходит любое вещество, непроницаемой — если она не пропускает ни одно вещество, избирательно проницаемой, или полупроницаемой, — если через нее могут диффундировать некоторые, но не все вещества. Все клеточные мембраны (окружающие саму клетку, ядра, вакуоли и различные субклеточные структуры) обладают дифференциальной проницаемостью.
Диффузия растворенного вещества через полупроницаемую мембрану называется диализом. Для того чтобы продемонстрировать процесс диализа, можно взять мешочек из коллодия, целлофана или пергамента, наполнить его раствором сахара и поместить в сосуд с водой. Если поры мембраны не слишком мелки, то молекулы сахара будут проходить сквозь нее. Со временем концентрация сахара в воде, окружающей мешочек, станет равна концентрации его в мешочке. Диффузия молекул будет продолжаться и после этого, но концентрация изменяться не будет, так как диффузия в обоих направлениях будет происходить с одинаковой скоростью.
Но если взять мешочек с более мелкими порами, с тем чтобы он был проницаем для молекул воды, но непроницаем для более крупных молекул сахара, то наблюдается другое явление. Нальем в мешочек раствор сахара, снабдим мешочек пробкой с проходящей через нее стеклянной трубкой и поместим его в сосуд с водой. Молекулы сахара не проходят через мембрану и поэтому остаются внутри мешочка. Молекулы воды, однако, диффундируют через мембрану в раствор сахара. Жидкость внутри мешочка содержит 5% сахара, а следовательно, лишь 95% воды. Жидкость же, окружающая мешочек снаружи, представляет собой чистую воду. Поэтому молекулы воды переходят из области более высокой концентрации (100% — снаружи) в область более низкой концентрации (95% — внутри мешочка). Такая диффузия молекул воды или другого растворителя через мембрану называется осмосом.
По мере того как происходит осмос, вода поднимается по стеклянной трубке. Если через мембраны пройдет такое количество воды, какое первоначально содержалось в мешочке, то раствор сахара окажется разбавленным до соотношения 2,5% сахара: 97,5% воды, но концентрация воды снаружи все еще будет выше, чем концентрация ее внутри, и осмос будет продолжаться. В конце концов уровень воды в стеклянной трубке поднимется настолько что давление, производимое водой в трубке, будет равно силе, заставляющей воду входить в мешочек. После этого изменение количества воды в мешочке прекратится; осмос через полупроницаемую мембрану будет продолжаться с одинаковой скоростью в обоих направлениях.
Давление столба воды в трубке служит мерой осмотического давления раствора сахара. Осмотическое давление вызывается стремлением молекул воды пройти сквозь полупроницаемую мембрану и выравнять концентрацию воды по обе стороны мембраны. Более концентрированный раствор сахара имел бы еще более высокое осмотическое давление и «насасывал» бы воду в трубку до более высокого уровня. При 10%-ной концентрации раствора сахара вода в трубке поднялась бы приблизительно вдвое выше, чем при 5%-ной.
Диализ и осмос — всего лишь две особые формы диффузии. Диффузия — общий термин для обозначения движения молекул из области высокой концентрации в область более низкой концентрации под действием тепловой энергии этих молекул. Диализом называется диффузия молекул растворенного вещества через полупроницаемую мембрану, а осмосом — диффузия через такую же мембрану молекул растворителя. В живых системах растворителем служит вода.
В жидком содержимом всякой живой клетки растворены соли, сахара и другие вещества, благодаря которым оно обладает известным осмотическим давлением. Если поместить клетку в жидкость с таким же осмотическим давлением, как и давление в клетке, вода не входит в клетку и не выходит из нее, а клетка соответственно не набухает и не съеживается; такую жидкость называют изотонической или изоосмотической по отношению к внутриклеточной жидкости. В норме плазма крови и все жидкости организма изотоничны; они содержат такое же количество растворенного материала, как и клетки.
Если концентрация растворенных веществ в окружающей жидкости выше, чем внутри клетки, то вода стремится выйти наружу и клетка съеживается. Такую жидкость называют гипертонической по отношению к клетке. Если в жидкости меньше растворенного материала, чем в клетке, ее называют гипотонической; в этом случае вода стремится входить в клетку и вызывает ее набухание. 0,9%-ный раствор хлористого натрия (иногда называемый «физиологическим» раствором) изотоничен по отношению к клеткам человека. Эритроциты, помещенные в 0,6%-ный раствор хлористого натрия, набухают и лопаются (рис. 21), в 1,3%-ном растворе они сжимаются, а в 0,9%-ном растворе с ними не происходит ни того, ни другого.
Если клетку поместить в раствор, не изотонический по отношению к ее содержимому, то она может иногда приспособиться к такой среде, изменяя содержание воды в своей протоплазме (набухая или сжимаясь) до тех пор, пока концентрация растворенных веществ в клетке и в окружающей среде не станет одинаковой. Многие клетки способны активно насасывать воду или некоторые растворенные вещества через плазматическую мембрану и откачивать их наружу, в результате чего поддерживается осмотическое давление, отличное от осмотического давления окружающей среды. У простейших, живущих в сильно гипотонической пресной воде, выработались сократительные вакуоли, которые вбирают в себя воду из протоплазмы и выводят ее наружу. Перед растениями, обитающими в пресной воде, тоже встает проблема: как быть с водой, проникающей в клетки из окружающей гипотонической среды путем осмоса? Клетки растений не имеют сократительных вакуолей, чтобы «откачивать» воду, но зато прочная целлюлозная стенка клетки предохраняет ее от чрезмерного набухания. По мере поступления в клетку воды в ней возникает внутреннее давление, называемое тургором, которое препятствует дальнейшему проникновению воды. Тургор вообще характерен для растительных клеток; именно он «поддерживает» тело растения. Когда тургорное давление в клетках понижается вследствие недостатка воды, цветок увядает.
Многие организмы, обитающие в море, обладают феноменальной способностью избирательно накапливать некоторые вещества из морской воды. Морские водоросли могут накапливать йод в таком количестве, что концентрация его в клетках становится в 2 миллиона раз выше, чем в окружающей среде. Примитивные хордовые — оболочники — способны накапливать ванадий, и концентрация этого элемента в их клетках может превышать его концентрацию в морской воде тоже примерно в два миллиона раз. Перенос воды или растворенных веществ в клетку или из клетки против градиента концентрации представляет собой физическую работу и требует затраты энергии. Клетка способна перемещать молекулы против градиента концентрации лишь пока она жива и пока в ней происходят метаболические процессы, доставляющие энергию. Если на клетку воздействовать каким-нибудь метаболическим ядом («отравляющим» обмен веществ), например цианистым калием, она утрачивает способность создавать и поддерживать разность концентраций по обе стороны плазматической мембраны.
КЛЕТОЧНОЕ ЯДРО
Каждая клетка содержит небольшую, обычно шаровидную или овальную органеллу, называемую ядром. В одних клетках ядро занимает относительно постоянное положение и располагается примерно в центре клетки; в других оно свободно перемещается и может оказаться почти в любом участке. Ядро играет важную роль в регулировании протекающих в клетке процессов; оно содержит наследственные факторы, или гены, определяющие признаки данной клетки и всего организма, и прямо или косвенно регулирует многие стороны клеточной активности. Ядро отделено от окружающей цитоплазмы ядерной мембраной, состоящей из двух элементарных мембран; ядерная мембрана регулирует передвижение веществ из ядра в цитоплазму и обратно. В электронном микроскопе видно, что ядерная мембрана состоит из двух слоев и что в ней имеются поры (рис. 22), через которые содержимое ядра сообщается с цитоплазмой; возможно, что через эти поры проходят информационные макромолекулы. По-видимому, наружный из двух слоев ядерной мембраны без перерыва переходит в мембраны эндоплазматической сети и комплекса Гольджи.
В ядре клетки, убитой путем фиксации в соответствующих химических веществах и окрашенной подходящими красителями, выявляются различные структуры. В живой клетке их трудно увидеть с помощью обычного светового микроскопа, но они хорошо видны в фазово-контрастном микроскопе (рис. 3). В кариоплазме — полужидком основном веществе ядра — взвешено строго определенное число вытянутых нитевидных образований, называемых хромосомами; они состоят из ДНК и белка и содержат единицы наследственности — гены. На окрашенном срезе (рис. 3) клетки в состоянии покоя (в период между делениями) хромосомы обычно неразличимы, а вместо них
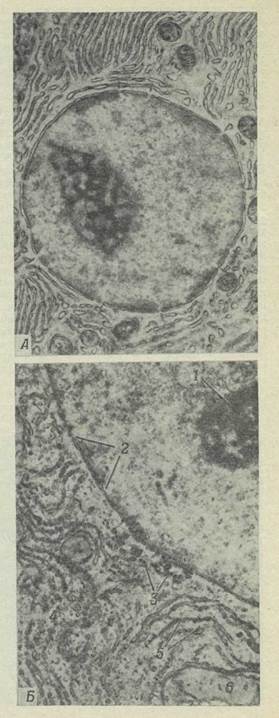 Рис. 22.
Рис. 22.
А. Электронная микрофотография ядра и окружающей его эндоплазматической сети; поры показаны стрелками (Х 20 000). Б. Часть того же препарата при большем увеличении (Х 50 000). 1 — ядрышко; 2 — поры; 3 — гликоген; 4 — рибосомы; 5 —эндоплазматическая сеть; е — митохондрия.
видна сеть из темных тяжей и зернышек, в совокупности называемых хроматином. Перед началом деления ядра эти тяжи уплотняются в компактные палочковидные хромосомы, которые впоследствии строго поровну распределяются между двумя дочерними клетками. Для каждого организма характерно строго определенное число хромосом, содержащееся в каждой из составляющих его клеток. У плодовой мушки (дрозофилы) 8 хромосом, у сорго их 10, у садового гороха 14, у кукурузы 20, у жабы 22, у томата 24, у вишни 32, у крысы 42, у человека 46, у картофеля 48, у козы 60, а у утки 80. Приведенные цифры относятся к соматическим клеткам высших растений и животных, в которых хромосома каждого типа представлена в двойном числе; клетку с двумя полными наборами хромосом называют диплоидной. Сперматозоиды и яйцеклетки, в которых каждая хромосома представлена лишь в единственном числе (т. е. имеется один полный набор хромосом), называют гаплоидными клетками. Число хромосом в них вдвое меньше, чем в соматических клетках того же организма. При оплодотворении яйцеклетки сперматозоидом два гаплоидных набора хромосом объединяются и таким образом восстанавливается их диплоидное число. В ядре имеется сферическое тельце, называемое ядрышком. В большинстве клеток ядрышко чрезвычайно изменчиво: оно меняет свою форму и структуру, появляется и исчезает. Ядро может содержать и несколько ядрышек, но обычно клетки каждого вида животных или растений содержат определенное число ядрышек. Ядрышки исчезают, когда клетка готовится к делению, а затем появляются вновь; они, по-видимому, участвуют в синтезе рибонуклеиновой кислоты, входящей в состав рибосом. Если ядрышко разрушить сфокусированным пучком рентгеновских или ультрафиолетовых лучей, то клеточное деление подавляется. Если же подвергнуть облучению другой участок ядра, не затрагивая ядрышко, то этого не происходит.
Для того чтобы выяснить роль ядра, можно удалить его из клетки и наблюдать последствия такой операции. Если с помощью микроиглы удалить ядро у одноклеточного животного — амебы, то клетка продолжает жить и двигаться, но не может расти и через несколько дней погибает. Следовательно, ядро необходимо для метаболических процессов (в первую очередь — для синтеза нуклеиновых кислот и белков), обеспечивающих рост и размножение клеток.
Можно возразить, что к гибели приводит не утрата ядра, а сама операция. Для того чтобы выяснить это, необходимо поставить опыт с контролем, т. е. подвергнуть две группы амеб одной и той же операции, с той разницей, что в одном случае ядро действительно удаляют, а в другом в амебу вводят микроиглу, передвигают ее в клетке подобно тому, как это делается при удалении ядра, и выводят, оставив ядро в клетке; это называется «мнимой» операцией. После такой процедуры амебы оправляются, растут и делятся; это показывает, что гибель амеб первой группы вызывалась не операцией как таковой, а именно удалением ядра.
Классическую серию опытов, доказывающих важную роль ядра в регуляции клеточного роста, провел Геммерлинг на одноклеточном (неклеточном) растении Acetahularia mediter-тапеа. Эта морская водоросль, которая может достигать высоты 5 см, внешне несколько напоминает гриб и имеет «корни» и «стебель», заканчивающийся вверху большой дисковидной «шляпкой». Все растение представляет собой одну-единственную клетку и содержит лишь одно ядро, находящееся у основания стебля. Геммерлинг установил, что если перерезать стебель, то нижняя часть остается живой, регенерирует шляпку и полностью оправляется после операции. Верхняя же часть, лишенная ядра, живет в течение некоторого времени, но в конце концов погибает, не будучи в состоянии восстановить нижнюю часть. Следовательно, ацетабулярии, так же как и амебе, ядро необходимо для метаболических процессов, лежащих в основе роста; регенерация — это, конечно, одна из форм роста. В последующих экспериментах Геммерлинг сначала перерезал стебель непосредственно над ядром, а затем вторично перерезал его чуть ниже шляпки. Изолированный отрезок стебля, будучи снова помещен в морскую воду, частично или полностью восстанавливал шляпку. Казалось бы, это говорит о том, что ядро не является необходимым для регенерации; однако если Геммерлинг удалял и вторую шляпку, то третья шляпка уже не развивалась.
На основании таких экспериментов Геммерлинг заключил, что ядро вырабатывает какое-то вещество, необходимое для образования шляпки. Это вещество распространяется путем диффузии вверх по стеблю и стимулирует рост шляпки. В только что описанных опытах после перерезок 1 и 2 в стебле оставалось достаточно этого вещества, чтобы вызвать образование еще одной шляпки. Однако после того, как это вещество было израсходовано на образование одной новой шляпки, регенерация второй шляпки при отсутствии ядра оказалась уже невозможной.
У другого вида, Acetabularia crenulata, шляпка ветвистая, а не дисковидная. Если кусочек стебля этого вида (без ядра) пересадить на нижнюю часть стебля A. mediterranea (с ядром A. mediterranea), то на верхушке стебля образуется новая шляпка, но форма ее определяется не пересаженным кусочком стебля, а нижней частью, на которую он пересажен. Ядро благодаря содержащимся в нем генам доставляет специфическую информацию, определяющую форму регенерирующей шляпки, и его влияние оказывается сильнее тенденции пересаженного кусочка стебля образовать шляпку, характерную для вида A. crenulata. Ядро может контролировать активность других частей клетки благодаря тому, что в его хромосомах закодированы «инструкции», необходимые для синтеза белков и других веществ, от которых зависят структурные особенности и функции клетки. Очевидно, всякий раз, когда клетка делится, весь этот набор инструкций удваивается и каждая из дочерних клеток получает по одному его экземпляру.