Bruusgaard J.C. Myonuclei acquired by overload exercise precede hypertrophy and are not lost on detraining
Bruusgaard, J.C. Myonuclei acquired by overload exercise precede hypertrophy and are not lost on detraining /J.C. Bruusgaard, I.B. Johansen, I.M. Egner, Z.A. Rana, and K. Gundersen// PNAS, V.104.–№34.– 2010.– P.15111-15116.
Миоядра приобретенные после силовой тренировки предшествуют гипертрофии и не теряются после прекращения тренировки
РЕЗЮМЕ
Эффект от предыдущих силовых тренировок может быть достаточно длительным, даже после большого перерыва в тренировках, что значительно облегчает последующую тренировку. Традиционно «мышечная память» была отнесена к нейронным факторам из-за отсутствия любых выявленных механизмов локальной памяти в мышечной ткани. Мы использовали методы визуализации для того, чтобы «вживую» изучать миоядра (ядра мышечных волокон), принадлежащие различным мышечных волокнам и заметили, что появление новые миоядер предшествует любому увеличению размеров мышц вследствие силовой тренировки с большими отягощениями. Старые и вновь приобретенные ядра сохраняются даже при сильной атрофии, возникающей при денервации в течение значительного периода жизни животного. Кажется, что миоядра защищены от апоптоза, возникающего в неактивной мышечной ткани. Гипертрофия, ведет к существенному повышению числа миоядер и тормозит проявления атрофии, а ядра могут служить в качестве субстрата для длительной биологической памяти клеток. В связи с тем, что способность увеличивать количество миоядер нарушается в пожилом возрасте, люди могут извлечь выгоду из силовых тренировок в раннем возрасте, и анаболические стероиды способствуют большему количеству миоядер, поэтому постоянство количества ядер может иметь последствия для исключения периодов после принятия допинга.
КЛЮЧЕВЫЕ СЛОВА: атрофия мышц, мышечная память, ядра мышечных волокон, гипертрофия мышц, апоптоз
ВВЕДЕНИЕ
Люди, занимающиеся силовой тренировкой очень быстро наращивают силу после ее прекращения и последующего возобновления (1, 2). Это явление уже окрестили «мышечной памятью». Так как до настоящего времени не был известен механизм памяти в мышечных волокнах, то на сегодняшний день этот феномен был отнесен к последствиям предыдущего моторного обучения в центральной нервной системе (3). Тем не менее, было сообщено, что мышцы могут оставаться гипертрофированными после нескольких месяцев прекращения тренировки (1, 4-8). В одном из исследований на пожилых людях, которые ранее тренировались, сила была еще на 9-14 % выше, даже после 2 лет отсутствия тренировок (7). Во время 30-32 недель отсутствия тренировок, группа женщин потеряла значительную часть дополнительной силы, полученной за 20 недель предыдущей тренировки, но восстановила уровень силы уже после 6 недель последующей тренировки (1). Это говорит о наличии локального механизма памяти в мышцах. Полученные данные объясняют этот феномен за счет изменения цитоархитектурных особенностей мышц после тренировки и гипертрофии, даже после последующего длительного бездействия.
Более чем 100 лет назад было высказано предположение, что ядро может поддерживать только определенный объем цитоплазмы (9-19). Мышечные клетки могут быть на пять порядков больше, чем одноядерные клетки и мышечные волокна являются одним из очень немногих типов многоядерных клеток у позвоночных (18). Когда мышечные волокна увеличиваются в размерах, предполагается, что увеличение количества ядер происходит за счет митоза и последующего слияния мышечных стволовых клеток и мышечных волокон, а при атрофии «избыток» миоядер удаляется селективным апоптозом некоторых ядер (20, 21). Такие механизмы должны были бы поддерживать постоянство размера цитоплазматического мионуклеарного домена. Последние данные с использованием методов покадровой визуализации в естественных условиях поставили под сомнение эту простую модель, потому что было показано, что во время атрофии сохраняется количество миоядер, соответствующее норме (22). Это становится возможным потому, что в гипертрофированных мышцах ядра, которые претерпели митоз, при бездействии склонны к апоптозу (23). Полученные данные показывают, что новые миоядра появившиеся после тренировок с большими отягощениями сохраняются не менее трех месяцев после денервации мышцы. Это изменение количества миоядер может быть основой «мышечной памяти».
РЕЗУЛЬТАТЫ
Появление новых миоядер предшествует гипертрофии
Мышца extensor digitorum longus (EDL) была перегружена за счет частичной ампутации основных синергистов (день 0), а ядра отдельных мышечных волокон были визуализированы у интактного животного путем введения меченых нуклеотидов на 1-21 дней после ампутации (рис. 1А). Количество ядер и площадь поперечного сечения (CSA) были определены при наблюдении в микроскоп дистальной части каждого волокна. Во время 21 -дневного периода, CSA увеличился на 35 % по сравнению с началом тренировок с 1474 ± 93 мкм2 до 1991 ± 150 мкм2, в то время как количество миоядер повысилось на 54% (с 41 ± 1,5 ядер/мм длины волокна до 63 ± 2,3 ядер/мм длина волокна), рис.1А. Количество ядер начало расти после шести дней и стабилизировалось после 11 дней. Это предшествовало увеличению поперечного сечения мышцы, которое начало увеличиваться только через девять дней и стабилизировалось через 14 дней (рис. 1B). Эта последовательность была подтверждена на отдельных волокнах путем расчета цитоплазматического объема, приходящегося на одно ядро для каждого волокна. Этот объем снизился на 16 % с 33749 ± 1457мкм3 в первые три дня (0-3 дней) после перегрузки до 28360 ± 956 мкм3 на 6-10 день после перегрузки (Р ≤ 0,01). Средний объем (31235 ± 1476 мкм3) оказался ниже, но не было статистически значимых отличий от контрольной группы на 12-21 день после перегрузки.
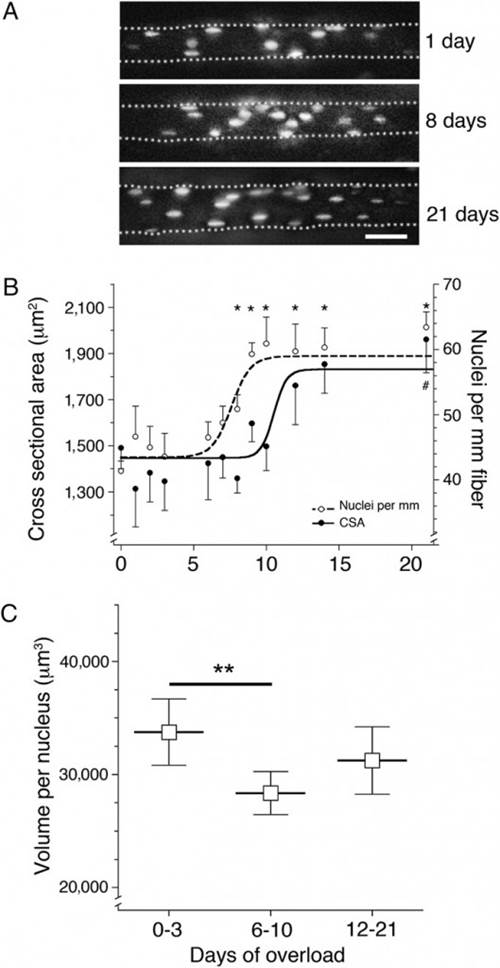
Рис 1. Влияние перегрузки на объем мышечного волокна и количество миоядер изученное в естественных условиях.
(A) Фотография мышечных волокон после перегрузки. Ядра помечены флуоресцентными олигонуклеотидами. Иллюстрации объединены в стеки изображений из различных фокальных плоскостей (Шкала:– 50 мкм).
(B) Кол-во ядер на миллиметр и площадь поперечного сечения (CSA). Каждая точка данных представляет 5-24 волокон от 36 животных. Символы представляют собой M ± m. Нелинейная зависимость в виде сигмовидной кривой, описывающей «доза-реакция» была использована для описания увеличения поперечного сечения (CSA) и количества ядер на миллиметр, в результате чего R2 =0,13 и R2 =0,29, соответственно. * Количество ядер на миллиметр CSA значительно отличается от начала эксперимента (день 0), P <0,05.
(C) Миоядерные домены. Каждый символ представляет собой время группы и рассчитывается от установленных в B данных **– статистические различия между измеренным временем групп (Р < 0,01).