В.Х. ХАВИНСОН
ПЕПТИДНАЯ РЕГУЛЯЦИЯ СТАРЕНИЯ

САНКТ-ПЕТЕРБУРГ
«НАУКА»
V.KH. KHAVINSON
PEPTIDE REGULATION OF AGEING
ST. PETERSBURG
«NAUKA»
2009
РОССИЙСКАЯ АКАДЕМИЯ НАУК
ГЕРОНТОЛОГИЧЕСКОЕ ОБЩЕСТВО
РОССИЙСКАЯ АКАДЕМИЯ МЕДИЦИНСКИХ НАУК
СЕВЕРО-ЗАПАДНОЕ ОТДЕЛЕНИЕ
САНКТ-ПЕТЕРБУРГСКИЙ ИНСТИТУТ БИОРЕГУЛЯЦИИ И ГЕРОНТОЛОГИИ
----------------------------------
RUSSIAN ACADEMY OF SCIENCES
GERONTOLOGYCAL SOCIETY
RUSSAN ACADEMY OF MEDICAL SCIENCES
NORTH-WESTERN BRANCH
SAINT PETERSBURG INSTITUTE OF BIOREGULATION AND GERONTOLOGY
В.Х. Хавинсон. Пептидная регуляция старения. – СПб.: Наука. 2009. с.
В научном издании обобщены результаты многолетних исследований автора и коллектива Санкт-Петербургского института биорегуляции и геронтологии СЗО РАМН, посвященных изучению механизмов старения и эффективности пептидных биорегуляторов в профилактике возрастной патологии. Приводятся данные по оценке действия пептидов, полученные с использованием наиболее современных методик в научных учреждениях России, США, Великобритании, Германии, Италии, Испании, Франции. Представлена молекулярная модель комплементарного взаимодействия коротких пептидов с промоторными участками генов, которая лежит в основе инициации синтеза белка. Предлагается концепция пептидной регуляции старения. Издание представляет интерес для широкого круга врачей, биологов, химиков.
Ответственный редактор:
академик РАН и РАМН А.И. Григорьев
Рецензенты:
академик РАН С.Г. Инге-Вечтомов,
академик РАН А.Д. Ноздрачев
V.Kh. Khavinson. Peptide regulation of ageing. – Saint Petersburg: Nauka. 2009 p.
The book generalizes the results of long-term author's investigation at the St. Petersburg Institute of Bioregulation and Gerontology of the North-Western Branch of the Russian Academy of Medical Sciences. It focuses on the mechanisms of ageing and geroprotective action of peptide bioregulators and highlights the data obtained by the most up-to-date methods at the research institutions in Russia, USA, Great Britain, Germany, Italy, Spain and France. The author outlined a molecular model of complementary interaction of small peptides with promoter section of genes which underlies initiation of protein synthesis. The book narrates a concept of peptide regulation of ageing, thus addressing a wide readership of physicians, biologists and chemists.
Научное издание
Утверждено к печати
Президиумом Геронтологического общества Российской академии наук и
Ученым Советом Санкт-Петербургского института биорегуляции и геронтологии СЗО РАМН
Содержание
1. Введение
2. Открытие пептидной регуляции старения
3. Заключение
4. Благодарность
5. Список литературы
Введение
В течение многих лет феномен старения рассматривался в рамках этических и социальных проблем. Только за последнее столетие общество осознало, что процесс старения нужно исследовать в другом аспекте: как специальный физиологический механизм организма, имеющий определённое эволюционное значение.
Старение – самая сложная проблема медицины и биологии. Процесс старения – это постепенная инволюция тканей и нарушение функций организма. Симптомы старости появляются уже в конце репродуктивного периода и становятся более интенсивными по мере дальнейшего старения.
В конце ХІХ века И.И. Мечников показал, что повышение клеточного иммунитета способствует увеличению продолжительности жизни. Он разработал фагоцитарную теорию иммунитета и считал, что в самом организме человека заложены возможности, позволяющие успешно бороться с патологической старостью [105]. В 1908 г. он был удостоен Нобелевской премии по физиологии или медицине совместно с П. Эрлихом. И только через столетие П. Догерти и Р. Цинкернагель выполнили детальные исследования специфичности клеточного иммунитета при вирусной инфекции (Нобелевская премия по физиологии или медицине в 1996 г.) [23].
Д. Уотсон и Ф. Крик совместно с М. Уилкинсоном получили Нобелевскую премию по физиологии или медицине в 1962 г. «за открытие молекулярной структуры нуклеиновых кислот и ее значение в передаче информации в живой материи».
В 1961 г. Ф. Жакоб и Ж. Моно предложили модель генетической регуляции белкового синтеза при участии низкомолекулярного лиганда, который вытесняет репрессор и вызывает аллостерический конформационный переход в структуре ДНК в бактериальной клетке [80]. Они получили Нобелевскую премию по физиологии или медицине в 1965 г. вместе с А. Львовым [23].
В результате многолетней работы М. Ниренберг и Г. Корана расшифровали генетический код и смогли определить кодоны (триплеты нуклеотидов) для каждой из двадцати аминокислот (Нобелевская премия по физиологии или медицине в 1968 г. совместно с Р. Холли) [23].
Фундаментальные исследования биохимии нуклеиновых кислот и определение последовательности оснований в РНК и ДНК были выполнены в 60 – 70 годы ХХ столетия П. Бергом, У. Гильбертом и Ф. Сэнджером (Нобелевская премия по химии в 1980 г.).
Экспериментальные и клинические исследования в геронтологии показали, что иммунная защита организма является первой системной функцией, которая нарушается при старении [68, 78]. Пептидные экстракты тимуса и пептиды, выделенные из этих экстрактов, были первыми препаратами, предложенными для коррекции иммунодефицитного состояния [67, 74, 76].
Происхождение пула коротких регуляторных пептидов в организме стало очевидным после открытия А. Чихановером, А. Гершко и И. Роузом убиквитин-опосредованной деградации белков в протеосомах (Нобелевская премия по химии в 2004 г.). В их работах было показано, что короткие пептиды играют важную роль в передаче биологической информации, как например, аутокринные гормоны и нейропептиды. Один высокомолекулярный белок может быть гидролизован различными путями, что приводит к возникновению нескольких коротких пептидов. Этот механизм может производить пептиды, несущие совершенно различные биологические функции по сравнению с исходной макромолекулой [79]. В работах американского математика С. Карлина было показано, что в макромолекулах белков имеется несколько типов повторяющихся блоков аминокислотных остатков с заряженными боковыми группами. Наибольшее количество таких блоков содержится в ядерных белках: факторах транскрипции, белках центромеров и группе белков высокой подвижности [81]. Протеосомный гидролиз этих белков в ядре может обеспечить присутствие достаточного набора пептидов с заряженными боковыми группами.
До начала работ коллектива нашего института регулирующая роль коротких пептидов в теориях генного контроля синтеза белков высших организмов не рассматривалась.
При старении, кроме снижения иммунитета, происходят и другие изменения на клеточном уровне. В частности, внутренняя структура клеточного ядра также изменяется в процессе старения. ДНК-белковый комплекс клеточного ядра (хроматин) самоорганизуется в хромосомы только при клеточном делении. В стационарном состоянии хроматин существует в двух разновидностях: эухроматин и гетерохроматин [103]. Гетерохроматин обычно локализован на периферии ядра и содержит в целом неактивную часть генома: гены, блокированные репрессорами. Соотношение эухроматин/гетерохроматин меняется при старении за счёт снижения содержания активного эухроматина, что определяет снижение синтеза белка в клетке [104].
Таким образом, старение организма имеет много уровней дисфункции и может быть классифицировано как системный синдром. Перспективные результаты коррекции иммунодефицитов с помощью эндогенных регуляторных пептидов указывали на необходимость дальнейшего расширения исследований [74, 76].
Открытие пептидной регуляции старения
Известно, что видовой предел продолжительности жизни животных и человека примерно на 30-40% превышает среднюю длительность жизни. Это связано с воздействием на организм различных неблагоприятных факторов, которые приводят к изменению экспрессии и структуры генов, что сопровождается нарушением синтеза белка и снижением функций организма (рис. 1) [1, 22, 35, 39].
Видовой предел жизни человека - 110-120 лет

Рис. 1. Видовая продолжительность жизни человека и его биологический резерв.
Современная медико-демографическая ситуация в России характеризуется высокой преждевременной смертностью, уменьшением рождаемости, снижением средней продолжительности жизни, что в сочетании с ростом числа лиц пожилого и старческого возраста ведет к депопуляции населения и дефициту трудового потенциала [1, 12, 90].
В последнее десятилетие достижения в теоретической и прикладной геронтологии позволили осуществлять целенаправленную регуляцию возрастных изменений. Исходя из этого, одной из приоритетных задач современной геронтологии является профилактика ускоренного старения и возрастной патологии, направленная на увеличение средней продолжительности жизни, сохранение активного долголетия и достижение видового предела жизни человека [35, 73, 77, 90].
Применение достижений фундаментальной науки в медицине привело к пониманию того, что прогресс клинической медицины во многом зависит от медицины молекулярной, т.е. исследований, проводимых на уровне генов и биологически активных молекул. Молекулярная медицина также широко использует достижения генетики, молекулярной и клеточной биологии для конструирования новых лекарственных средств и технологий [1, 24, 73].
Одним из актуальных направлений молекулярной медицины является изучение генетических механизмов старения. В настоящее время установлено, что существуют гены, которые регулируют механизмы индивидуального развития и возникновения многих заболеваний [7, 8, 99].
При возрастном снижении процессов пролиферации и дифференцировки клеток существует возможность осуществлять коррекцию этих нарушений путем воздействия на экспрессию генов [20, 72, 73, 89]. Изучение генетических механизмов старения и развития возрастной патологии составляет основу регуляторной терапии – использования модуляторов транскрипции, сдерживающих и восстанавливающих наступающие с возрастом генетические изменения. Для этого необходимо знание генома, возникающих нарушений и использование веществ селективного воздействия на экспрессию генов [29, 36, 89]. Создание эффективных биорегуляторов, способствующих достижению видового предела продолжительности жизни и сохранению основных физиологических функций, является одной из наиболее актуальных проблем современной биогеронтологии. В исследованиях, посвященных данной проблеме, значительное внимание уделяется роли пептидов в предотвращении ускоренного старения [35, 82, 89].
Пептидная регуляция гомеостаза занимает важное место в сложной цепи физиологических процессов, приводящих к старению клеток, тканей, органов и организма в целом. Морфо-функциональным эквивалентом старения является инволюция органов и тканей и прежде всего тех, которые относятся к основным регуляторным системам – нервной, эндокринной и иммунной [16, 25, 37]. Имеются данные, свидетельствующие о возрастной гипоплазии, а в ряде случаев и об атрофии шишковидной железы (эпифиза), тимуса, нейронов коры головного мозга и подкорковых структур, сетчатки, сосудистой стенки, половых органов [10, 27, 37, 66].
В начале 1970-х гг. мы изучали механизм иммунодепрессии в эксперименте и клинике. Было установлено, что по мере старения происходит инволюция центрального органа иммунной системы - тимуса (рис. 2, 3) и нейроэндокринной системы – эпифиза [11, 21, 35]. Также было выявлено достоверное снижение синтеза белка в клетках различных тканей организма (рис. 4) [22, 35].

| Субкапсулярная зона коры (ребенок 2 года) А - окраска гематоксилином и эозином; В - свечение полипептидов тимуса в телах и отростках, образующих ячейки Кларка, а также в виде гранул на мембранах тимоцитов внутри ячеек. |

| Субкапсулярная зона коры (мужчина 46 лет) А - окраска гематоксилином и эозином; В - свечение полипептидов тимуса в телах и отростках эпителиальных клеток, образующих группы по 2-5 клеток. |
Рис. 2. Возрастная инволюция тимуса (непрямой иммунофлюоресцентный метод с антителами к полипептидам тимуса, х600).

|
Иммунофлюоресцентная лазерная конфокальная микроскопия, х400 (красное свечение - Rodamin G, зеленое свечение - FITC).
| |||||||||||||

|
Рис. 3. Синтез транскрипционных протеинов (PAX 1) в тимусных эпителиальных клетках человека (исследование выполнено совместно с Биомедицинским научно-исследовательским центром принца Филиппа, Валенсия, Испания).

*- р<0.05 по сравнению с показателем у крыс 3 мес;
**- p<0.05 по сравнению с показателем у крыс 9 мес.
Рис. 4. Синтез белка в гепатоцитах крыс разного возраста.
Для восстановления функций тимуса, эпифиза, костного мозга и других органов нами был разработан специальный метод выделения и фракционирования низкомолекулярных пептидов из экстрактов этих органов [18, 43, 69, 106, 109].
На уровне целого организма у различных животных было продемонстрировано значительное разнообразие биологической активности коротких пептидов и особенно пептидного препарата тимуса (лекарственный препарат «тималин») и препарата эпифиза (лекарственный препарат «эпиталамин») [5, 6, 17, 21, 22, 35, 65, 82, 92]. Эти пептидные препараты в многочисленных экспериментах способствовали достоверному увеличению средней продолжительности жизни животных до 25-30% по сравнению с контролем [2, 3, 4, 35, 52, 56, 63, 64]. В большинстве экспериментов отмечено также некоторое увеличение максимальной продолжительности жизни. Наиболее значимый эффект увеличения максимальной продолжительности жизни был отмечен у мышей CBA при введении им пептида Ala-Glu-Asp-Gly и составил 42,3% [55]. Особенно следует отметить отчетливую корреляцию увеличения средней продолжительности жизни и основного показателя клеточного иммунитета – реакции бласттрансформации лимфоцитов с фитогемагглютинином (РБТЛ с ФГА), характеризующего функцию Т-лимфоцитов, при введении препаратов тимуса и эпифиза животным (рис. 5) [22, 56].

* - p<0.01 по сравнению с контролем (0)
Рис. 5. Влияние пептидных препаратов на среднюю продолжительность жизни и РБТЛ с ФГА у мышей.
Значительное увеличение средней продолжительности жизни животных безусловно было связано с тем, что низкомолекулярные пептиды, выделенные из эпифиза и тимуса, обладали достоверной противоопухолевой активностью, что выражалось в резком снижении в 1,4-7 раз частоты возникновения как спонтанных, так и индуцированных облучением или канцерогенами злокачественных опухолей у животных (рис.6) [35, 49, 56, 57, 60, 62, 63, 70]. Следует подчеркнуть, что этот беспрецедентный уровень уменьшения количества опухолей был отмечен в подавляющем большинстве экспериментов (более 30). Результаты этих исследований, учитывая общий механизм канцерогенеза у всех млекопитающих, имеют огромное практическое значение для профилактики опухолей у людей [35, 57, 91].

* - p<0.05 по сравнению с контролем
Рис. 6. Влияние пептидного препарата эпифиза на частоту возникновения опухолей у животных.
В специальных экспериментах было установлено, что короткие пептиды, выделенные из различных органов и тканей, а также их синтезированные аналоги (ди-, три-, тетрапептиды) обладают выраженной ткане-специфической активностью как в культуре клеток, так и в экспериментальных моделях у молодых и старых животных (рис. 7) [22, 36, 82].
Воздействие пептидов приводило к тканеспецифической стимуляции синтеза белка в клетках тех органов, из которых эти пептиды были выделены. Эффект усиления синтеза белка при введении пептидов выявлен у молодых и старых животных (рис. 8).

* - p<0.05 по сравнению с контролем (0)
Рис. 7. Пептидная тканеспецифическая регуляция роста эксплантатов тканей в органотипических культурах клеток.
|

* - p<0.05 по сравнению с контролем
Рис. 8. Влияние пептидов на синтез белка в гепатоцитах крыс разного возраста.
Особенно значимым явился факт восстановления репродуктивной системы у старых самок крыс после введения пептидного препарата эпифиза [6, 69]. Так, фаза эструса у животных, аналогичная менопаузе у женщин, от исходного состояния 95% уменьшилась после введения препарата до 52%, а остальные фазы цикла, характерные для нормы, возросли от исходных 5% до 48%. Необходимо подчеркнуть, что в другом эксперименте ни у одной старой крысы беременность после спаривания с молодыми самцами не наступала. После введения препарата эпифиза при повторном спаривании у 4 крыс из 16 наступила беременность, и у них родились по 5-9 здоровых крысят.
Таким образом, были установлены главные преимущества низкомолекулярных пептидов по сравнению с высокомолекулярными белковыми регуляторами: они обладают высокой биологической активностью, проявляют тканеспецифичность, у них отсутствуют видоспецифичность и иммуногенность. Эти характеристики сближают регуляторные пептиды с пептидными гормонами [17, 21, 79, 102].
В течение многих лет проводилось подробное изучение молекулярных масс, химических свойств, аминокислотного состава и последовательности аминокислот низкомолекулярных пептидов из тимуса, эпифиза и других органов [21, 22, 106, 107, 108, 109]. Полученная информация была использована для осуществления химического синтеза некоторых коротких пептидов. Сравнение показало, что биологическая активность природных и синтетических препаратов в основном идентична. Так, например, дипептид тимуса GluTrp стимулировал иммунитет, снижал темп старения и подавлял возникновение спонтанных опухолей у животных [17, 58, 59, 108]. Биологическая активность природных и синтетических пептидов была сходной при стандартном тестировании в культуре тканей и у животных [21, 35, 36, 53, 71, 86, 101, 108]. Эти результаты указывали на перспективность применения пептидов в качестве геропротекторных препаратов [2, 3, 4, 83]. Учитывая актуальность поиска новых лекарственных средств – геропротекторов, были проведены доклинические исследования пептидных препаратов на различных уровнях.
На уровне клеточных структур было обнаружено, что короткие пептиды активируют гетерохроматин в клеточных ядрах людей старческого возраста и способствуют «высвобождению» генов, репрессированных в результате гетерохроматинизации эухроматиновых районов хромосом, которая происходит при старении (табл. 1) [88, 104].
Таблица 1
Влияние пептидов на хроматин в лимфоцитах лиц старческого возраста
| Пептиды | Ассоциирующие акроцентрические хромосомы (на клетку) | Дегетеро-хроматинизация факультативного гетерохроматина (на клетку) | Общий гетеро-хроматин | Структурный гетерохроматин 1 хромосомы |
| Контроль (20-40 лет) | 1.33±0.06 | 7.7±0.4 | стабильное состояние | стабильное состояние |
| Контроль (75-88 лет) | 1.17±0.05* | 5.9±0.2* | гетерохрома-тинизация | гетерохрома-тинизация |
| Пептид Lys-Glu | 2.39±0.11** | 9.9±0.6** | дегетерохро-матинизация | гетерохрома-тинизация |
| Пептид Ala-Glu-Asp-Gly | 2.32±0.12** | 8.4±0.5** | дегетерохро-матинизация | дегетерохро-матинизация |
* - p<0.05 по сравнению с контролем (20-40 лет)
** - p<0.001 по сравнению с контролем (75-88 лет)
Структурная конденсация хроматина находится в тесной корреляции с функциональной гетерогенностью. Установлено, что при старении усиливается гетерохроматинизация, которая коррелирует с инактивацией ранее активных генов [104]. Плотно конденсированные гетерохроматиновые районы хромосом генетически инактивированы, поздно реплицируются. Деконденсированные (эухроматиновые) районы хромосом активно функционируют. Известно, что необходимым условием для транскрипционной активности генов является активный хроматин [103]. Как уже упоминалось выше, в клеточном ядре существует две разновидности хроматина: светлый эухроматин и плотный гетерохроматин, расположенный рядом с ядерной мембраной. Транскрипция генов происходит в светлой фазе – в эухроматине. При старении объём гетерохроматина в ядре увеличивается в среднем от 63% до 80%. Регуляторные пептиды увеличивают содержание эухроматина в ядре. Это означает, что большее число генов оказывается доступным для факторов транскрипции, а транскрипция происходит более интенсивно, и синтез белка увеличивается. Иными словами, чем выше содержание эухроматина в ядре, тем интенсивнее синтез белка в клетке [88, 103, 104]. Результаты этого эксперимента позволили сделать крайне важный вывод о том, что гетерохроматинизация хроматина является обратимым процессом, а это подтверждает возможность восстановления синтеза белка и следовательно функций организма [36, 89, 104].
Важнейшим экспериментальным фактом явилось обнаружение способности пептидов индуцировать дифференцировку полипотентных клеток (рис. 9) [36, 38]. Так, добавление пептидов сетчатки к полипотентным клеткам эктодермы ранней гаструлы лягушки Xenopus laevis привело к возникновению клеток сетчатки и пигментного эпителия. Этот выдающийся результат в значительной степени объясняет положительный клинический эффект после применения препарата сетчатки у людей при дегенеративных заболеваниях сетчатки [27, 35, 82] и у животных с генетически детерминированным пигментным ретинитом [35, 94].

Рис. 9. Индукционное влияние пептидов сетчатки на полипотентные клетки эктодермы ранней гаструлы Xenopus laevis.
Добавление других коротких пептидов к полипотентным клеткам эктодермы в этой же экспериментальной модели приводило к возникновению различных тканей. Эти эксперименты показали, что пептиды способны индуцировать дифференцировку клеток в зависимости от структуры добавляемого вещества. Анализ результатов этих исследований дает основание сделать принципиальный вывод о возможности целенаправленной индукции дифференцировки полипотентных клеток и использования биологического клеточного резерва различных органов и тканей организма, что составляет основу увеличения продолжительности жизни до видового предела.
Известно, что число хромосомных аберраций используется как маркер повреждений ДНК в стареющем организме. Соматические мутации могут возникать из-за накопления устойчивых аберраций и лежат в основе возрастной патологии, включая злокачественные опухоли [111]. Достоверная антимутагенная и репаративная активность пептидов тимуса и эпифиза была подтверждена снижением числа хромосомных аберраций в клетках костного мозга и эпителия роговицы животных с ускоренным старением [36, 61, 89, 100].
На уровне регуляции активности генов установлено, что пептиды Lys-Glu и Ala-Glu-Asp-Gly при введении в организм трансгенных мышей подавляют экспрессию гена HER-2/neu (рак молочной железы человека в 2 – 3,6 раза по сравнению с контролем). Это подавление экспрессии гена сопровождается достоверным уменьшением диаметра опухоли (рис. 10) [35, 36, 62, 89].

*- р<0.001 по сравнению с показателем в контроле
**- р<0.05 по сравнению с показателем в контроле
Рис. 10. Влияние пептидов на развитие аденокарцином молочной железы и экспрессию онкогена HER-2/neu у трансгенных мышей (исследование выполнено совместно с Национальным центром старения, Анкона, Италия).
Обнаружено, что добавление пептида Ala-Glu-Asp-Gly в культуру легочных фибробластов человека и инкубирование их при 30º С в течение 30 мин индуцирует экспрессию гена теломеразы, активность теломеразы и способствует удлинению теломер в 2,4 раза. Активация экспрессии гена сопровождается увеличением числа делений клеток на 42,5%, что демонстрирует преодоление предела клеточного деления Хейфлика (рис. 11) [36, 89]. Этот важнейший результат полностью коррелирует с ранее указанным максимальным увеличением продолжительности жизни у животных (42,3%) после введения этого пептида [22, 55].
С использованием ДНК-микрочиповой технологии выполнено исследование влияния пептидов Lys-Glu, Glu-Trp, Ala-Glu-Asp-Gly, Ala-Glu-Asp-Pro на экспрессию 15247 генов сердца и головного мозга мышей [51, 89]. В экспериментах использовали клоны, входящие в библиотеку кДНК Национального института старения США. В этих экспериментах были получены уникальные данные по изменению экспрессии различных генов под влиянием пептидов (рис. 12). Важным выводом явилось то, что каждый пептид специфически регулирует конкретные гены. Результаты эксперимента указывают на существующий механизм пептидной регуляции генетической активности. В эксперименте было также установлено что дипептид Lys-Glu, обладающий иммуномодулирующей активностью, регулирует экспрессию гена интерлейкина-2 в лимфоцитах крови [36, 89].

Увеличение числа делений клеток в опыте по сравнению с контролем на 42.5%
* - р<0.05 по сравнению с контролем
Рис. 11. Преодоление лимита деления соматических клеток человека при добавлении пептида Ala-Glu-Asp-Gly в культуру легочных фибробластов.

Исследовано 15247 генов
(ДНК-микрочиповая технология)
(максимальное повышение в 6,61 раза, максимальное снижение в 2,71 раза)
Рис. 12. Влияние пептидов на экспрессию генов в сердце мыши (исследование выполнено совместно с Национальным институтом старения, Балтимор, США).
На молекулярном уровне существовал очевидный разрыв между многочисленными доказательствами специфических эффектов, вызванных регуляторными пептидами в активации транскрипции генов [36, 87, 89, 110], и ограниченными схемами процесса, который лежит в основе селективного связывания факторов транскрипции со специфическими сайтами ДНК. При этом неспецифическое связывание белков с двойной спиралью ДНК было доказано физико-химическими методами [45, 47]. Для активации транскрипции гена в клетках высших организмов, как правило, требуются десятки макромолекулярных активаторов и факторов транскрипции.
Нами была предложена молекулярная модель взаимодействия регуляторных пептидов и двойной спирали ДНК на промоторном участке гена (рис. 13, 14, 15, 16) [36, 45, 46, 89, 95].

Рис. 13. Развернутая конформация пептида Ala-Glu-Asp-Gly (проекция на плоскость). Представлены концевые и боковые функциональные группы, способные к комплементарным взаимодействиям с ДНК.
—NH3, —OH - протон-донорные группы;
=O - протон-акцепторные группы;
—CH3 - гидрофобная (метильная) группа.
Утолщенная линия обозначает основную пептидную цепь.
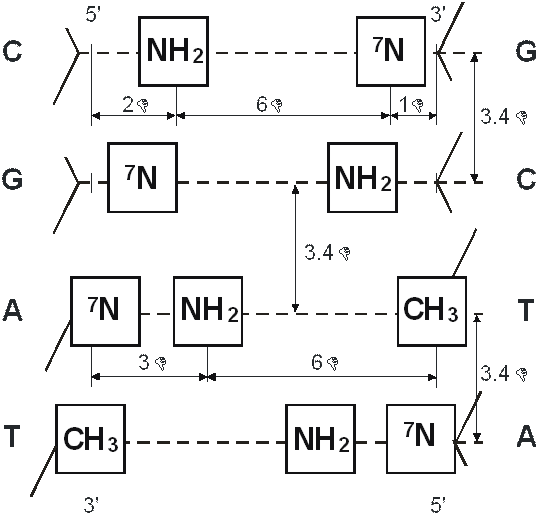
Рис. 14. Метрическое расположение функциональных групп на поверхности большой канавки при встраивании каждой нуклеотидной пары в двойную спираль ДНК.
Штриховая линия представляет перпендикулярную плоскость, в которой расположены ароматические структуры нуклеиновых оснований.
—NH2 - протон-донорные группы;
=7N - протон-акцепторные группы;
—CH3 - гидрофобная (метильная) группа.

Рис. 15. Последовательность нуклеотидных пар в двойной спирали ДНК, функциональные группы которой комплементарны функциональным группам пептида Ala-Glu-Asp-Gly.
Эта последовательность нуклеотидных пар многократно повторяется на промоторном участке гена теломеразы.

Рис. 16. Модель комплементарного взаимодействия пептида Ala-Glu-Asp-Gly с двойной спиралью ДНК (комплекс ДНК-пептид на промоторном участке гена теломеразы).
Геометрическая и химическая комплементарность аминокислотной последовательности пептида и последовательности нуклеотидных пар ДНК были положены в основу молекулярной модели. Регуляторный пептид распознаёт специфический сайт в двойной спирали ДНК, если его собственная аминокислотная последовательность комплементарна на достаточном протяжении последовательности нуклеотидов ДНК; другими словами – их взаимодействие специфично из-за совпадения последовательностей.
Каждая последовательность нуклеотидных пар в двойной спирали ДНК образует уникальный узор функциональных групп на поверхности большой канавки двойной спирали ДНК. Пептид в развёрнутой β-конформации может комплементарно расположиться в большой канавке ДНК вдоль оси двойной спирали. Литературные данные о молекулярной геометрии двойной спирали ДНК и пептидной β-нити были использованы для того, чтобы найти последовательность нуклеотидных пар для специфического связывания ДНК и пептида Ala-Glu-Asp-Gly. Скрининг показал, что этот тетрапептид может быть размещён в большой канавке ДНК с последовательностью нуклеотидов на ведущей цепи ATTTG (или ATTTC) в соответствии с комплементарностью расположения их функциональных групп [45, 47].
Для экспериментальной проверки молекулярной модели были использованы синтетические препараты: ДНК [поли(dA-dT):поли(dA-dT)] (двойная спираль) и пептидAla-Glu-Asp-Gly. С помощью гельхроматографии было доказано, что пептид Ala-Glu-Asp-Gly образует устойчивый межмолекулярный комплекс с двойной спиралью ДНК (рис. 17) [45, 89, 95].

1 - индивидуальный пептид Ala-Glu-Asp-Gly
2 - свободная двойная спираль ДНК [поли(dA-dT):поли(dA-dT)]
3 - смесь пептида и ДНК
Рис. 17. Гельхроматография пептида и ДНК на сефадексе G-25 в физиологическом растворе при комнатной температуре.
Комплементарное связывание пептида с последовательностью нуклеотидов на ведущей цепи ТАТАТА двойной спирали может быть осуществлено посредством шести водородных и одной гидрофобной связей между функциональными группами обоих участников.
В нормальных физиологических условиях ДНК существует в форме двойной спирали, две полимерных цепи которой удерживаются вместе водородными связями между парами оснований на каждой цепи. Большинство биологических процессов, включающих ДНК (транскрипция, репликация), требует, чтобы двойная спираль разделилась на отдельные цепи. В частности, известно, что локальное разделение цепей двойной спирали предшествует транскрипции генов РНК полимеразой. Для того, чтобы началась транскрипция (синтез матричной РНК), двойная спираль ДНК должна быть освобождена от гистонов, а в том месте, где начинается синтез матричной РНК, цепи двойной спирали должны быть разделены(рис. 18).

Рис. 18. Схема локального разделения цепей [поли(dA-dT):поли(dA-dT)] в результате связывания пептида Ala-Glu-Asp-Gly в большой канавке двойной спирали ДНК.
C использованием спектрофотометрии в ультрафиолетовой области растворов синтетической двойной спирали ДНК и пептида Ala‑Glu‑Asp‑Gly обнаружен концентрационно зависимый гиперхромный эффект (увеличение оптической плотности раствора при длине волны 260 нм) в смеси пептида и двуспиральной ДНК. Гиперхромный эффект свидетельствует о частичном разрушении водородных связей между нуклеотидными парами двойной спирали и о локальном разделении цепей двойной спирали (аллостерическое конформационное изменение) [44, 45].
В специальном эксперименте установлено, что разделение цепей (плавление) свободной синтетической ДНК происходит при температуре +69,50 С. В системе ДНК с пептидом плавление спирали произошло при +280 С и характеризовалось снижением показателей энтропии и энтальпии процесса примерно в 2 раза [44]. Этот важный факт указывает на практическую возможность термодинамически облегченного пути разделения цепей ДНК при температурном режиме, характерном для биохимических реакций большинства живых организмов. Это свидетельствует также о том, что разделение цепей ДНК при физиологической температуре не является денатурацией и характерно для инициации процесса синтеза белка. Эксперименты in vitro показывают, что короткий пептид определённой структуры и аминокислотной последовательности можетучаствовать в активации транскрипциигенов на этапе разделения цепей двойной спирали ДНК. Биохимический аспект этого факта состоит в сходстве структуры и аминокислотной последовательности регуляторного пептида и специфического участка пептидной цепи макромолекулярного фактора транскрипции.
Следует сделать выводы, что изучение биологической активности пептидов на различных структурных уровнях и исследование физико-химических процессов их взаимодействия показало несомненную высокую физиологическую активность пептидных регуляторов и перспективность их дальнейшего использования. Основным выводом явилось то, что пептиды обладают способностью регулировать экспрессию генов. В доклинических исследованиях установлена высокая биологическая активность и безопасность синтезированных пептидов [17, 19, 21, 35, 82]. Так, введение пептидов Lys-Glu, Ala-Glu-Asp-Gly животным способствовало уменьшению частоты развития опухолей и увеличению средней продолжительности жизни [9, 33, 34, 60, 61, 62]. Пептид Ala-Glu-Asp-Pro стимулировал регенерацию нерва [35, 36, 93], пептид Lys-Glu-Asp-Trp снижал уровень глюкозы в крови у животных с экспериментальным сахарным диабетом [32], пептид Ala-Glu-Asp увеличивал плотность костной ткани [26], пептид Ala-Glu-Asp-Leu способствовал восстановлению функций клеток бронхиального эпителия [97], пептид Ala-Glu-Asp-Arg восстанавливал функциональную активность клеток миокарда [98].
В настоящее время продолжается исследование пептидных препаратов, выделенных из хрящей, семенников, печени, сосудов, мочевого пузыря, щитовидной железы, а также синтезированных пептидов, регулирующих функцию мозга, сетчатки, иммунной системы, пролиферацию и дифференцировку полипотентных клеток. Эти физиологически активные вещества обладают, как правило, значительной тканеспецифической активностью и безусловно перспективны для создания на их основе новых лекарственных препаратов для биорегулирующей терапии [30, 31, 35].
Применение пептидных биорегуляторов у обезьян. Учитывая значительную достоверную биологическую активность пептидов, следующим целесообразным этапом явилось изучение пептидных регуляторов у обезьян (макак резусов, Macaca mulatta) [11, 75]. Важным достижением оказался результат полного восстановления уровня секреции мелатонина до нормы молодых животных (6-8 лет) у старых обезьян (20-26 лет) после введения пептида эпифиза (рис. 19) [11, 85].

* - p<0.01 по сравнению с контрольной группой молодых животных (плацебо)
**- p<0.001 по сравнению с контрольной группой старых животных (плацебо)
Рис. 19. Влияние пептида эпифиза на продукцию мелатонина у обезьян различного возраста.
У этих же старых обезьян после введения пептида восстановился до нормы суточный ритм секреции основного гормона надпочечников – кортизола (рис. 20). Введение пептида или препарата эпифиза старым животным привело также к восстановлению нарушающейся при старении толерантности к глюкозе. В