Силовой дефицит зависит от:
1) эмоционального (психологического) состояния человека (его настрой);
2) числа активных ДЕ (особенно больших);
3) совершенствования управления двигательными единицами центральной нервной системой.
Каждая двигательная единица имеет свой индивидуальный порог возбуждения, который может быть низким или высоким. Если импульсный залп (раздражение нерва, вызывающее сокращение мышцы) слаб, то тогда активизируются лишь двигательные единицы, обладающие низким порогом возбуждения. Если импульсный залп усиливается, начинают реагировать дополнительные двигательные единицы с более высоким порогом возбуждения
(рис. 2.9).
 |
Рис. 2.9. Зависимость силы произвольного сокращения от степени
рекрутирования мышечных волокон
С увеличением сопротивления активизируется все больше двигательных единиц. Быстрота индивидуальных порогов возбуждения зависит главным образом от состояния двигательных единиц. Для продолжения деятельности двигательных единиц, которые утомляются от: а ) накопления кислых продуктов обмена веществ (лактат, СО2); б) истощения энергоносителей (энергетические фосфаты, гликоген и т. п.); в) нервного перевозбуждения (в двигательной единице или в коре головного мозга), требуется все больше и больше волевых усилий.
Величина максимального мышечного напряжения зависит от центральных и периферических факторов.
К периферическим (структурным, внутримышечным) факторам относят:
1) количество мышечных волокон в мышце,
2) величину двигательных единиц, степень рекрутирования (вовлечения в сокращение) мышечных волокон.
3) длину мышечных волокон. Существует некоторая средняя длина Л0 (это длина мышцы при покое в условиях целостного организма), при которой мышца развивает максимальное сокращение. Если длина будет меньше Л0, или, наоборот, больше Л0 (перерастянута), то сила, развиваемая мышцей в момент ее возбуждения, будет значительно меньше. Оказалось, что максимальная сила развивается мышцей в том случае, когда длина саркомера составляет 2,2–2,5 мкм. Зависимость силы мышцы от ее длины очень важна, особенно для сердечной мышцы (закон Франка-Старлинга) в практическом и теоретическом отношениях (она доказывает гипотезу скольжения протофибрилл, объясняющую механизм сокращения);
4) тип строения мышцы (степень наклона мышечных волокон к оси движения – физиологический поперечник мышцы),
5) композицию мышцы (соотношение гликолитических, оксидативных и промежуточных волокон),
6) функциональных ( энергетический потенциал – содержание в мышце АТФ, КрФ, гликогена, миоглобина и сократительных белков).
К центральным факторам относят:
1) факторы внутримышечной координации (частота и характер нервных импульсов в ДЕ).
В 1885 году Н. Е. Введенский ввел понятие оптимума и пессимума частоты и силы раздражения, т. е. зависимости амплитуды ответной реакции мышцы от частоты и силы раздражения. Например, импульсы с частотой 30 Гц (30 имп/с) вызывают тетанус высотой 10 мм миографической записи,
50 Гц–15 мм, 200 Гц – 3 мм. В этом примере 50 Гц – оптимальная частота (оптимум), 200 Гц – пессимальная частота (пессимум). Таким образом, меняя частоту посылки импульсов к мышечным волокнам, альфа-мотонейрон может регулировать величину сократительного ответа своего мышечного аппарата.
К факторам внутримышечной координации также относится регуляция числа активных, возбуждаемых в данный момент времени ДЕ (Хенеманн). Так, если мышца представлена 10 ДЕ, а в данный момент активна 1ДБ, то мышца способна развить силу, равную 1/10 от ее максимальной силы. Если
5 ДЕ активны, то соответственно, мышца развивает 50% от максимума и т. д., а 100% силы она разовьет в том случае, если все 10 ДЕ одновременно будут возбуждены.
Так, если все 10 ДЕ начнут одновременно возбуждаться, то сила будет, например, 4 кгс/см2, а если они возбуждаются асинхронно, то максимальная сила составит 3 кгс/см2.
2) факторы межмышечной координации (мобилизация агонистов, торможение антагонистов, адаптационно-трофические влияния симпатической нервной системы – феномен Орбели-Гинецинского).
3) гормональные влияния (гормоны с анаболическим эффектом: половые, гормон роста и др.)
Для практических нужд важным является непосредственное измерение силовых показателей различных мышечных групп.
К методам измерения мышечной силы относят:
1) динамометрию;
2) динамографию;
3) электростимуляцию;
4) важными характеристиками силовых способностей являются время напряжения и расслабления, а также латентное время напряжения и расслабления.
2.4. Теория мышечного сокращения. Одиночное
и тетаническое сокращение мышц. Теория тетануса.
Формы и типы мышечных сокращений
Для понимания того, как происходит мышечное сокращение, необходимо рассмотреть микроскопическое строение мышечного волокна
(рис. 2.10). Скелетные мышцы состоят из мышечных пучков, образованных большим количеством мышечных волокон. Каждое волокно – это клетка цилиндрической формы диаметром 10–100 мкм и длиной от 5 до 400 мкм. Оно имеет клеточную мембрану – сарколемму. В саркоплазме находится несколько ядер, митохондрии (30–35% от массы волокна), образования саркоплазматического ретикулума (СР) и сократительные элементы – миофибриллы. Миофибриллы составляют в совокупности приблизительно 50% массы волокна, их длина равна длине мышечных волокон, и они, собственно, являются сократительными элементами мышцы.
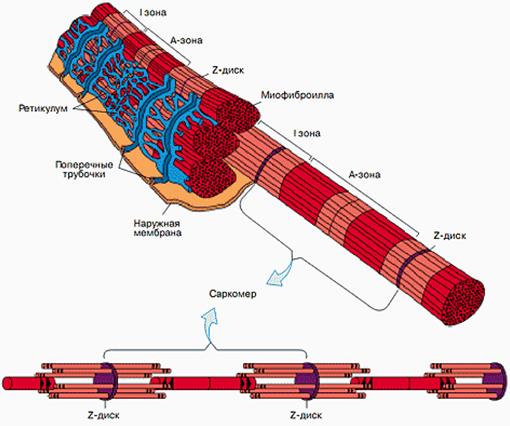 |
Рис. 2.10. Схема строения мышечного волокна [4]
Саркоплазматический ретикулум состоит из системы поперечных, продольных трубочек и цистерн. Поперечные трубочки – это впячивания сарколеммы внутрь клетки. К ним примыкают продольные трубочки с цистернами. Образования саркоплазматического ретикулума необходимы для распространения потенциала действия от сарколеммы на собственно мышечные волокна.
|
Темные полосы называют А-дисками или анизотропными ( миозиновые), обладающие анизотропией и поэтому имеющие темный цвет. I-диски (светлые полосы) – изотропные, образованы нитями актина. В центре I-дисков видна тонкая Z-пластинка. К ней прикрепляются актиновые протофибриллы.
Участок миофибриллы между двумя Z-пластинками называется саркомером. Саркомер – структурный элемент миофибрилл.
Рис. 2.11. Взаимное расположение волокон актина и миозина
в различных участках саркомера.
 |
Белки актин и миозин образуют основные сократительные элементы мышц – актомиозиновые комплексы саркомеров. Миозиновые и актиновые филаменты расположены параллельно друг другу строго определенным образом: в каждом участке миофибриллы вокруг толстого миозинового филамента расположено шесть тонких актиновых филаментов (рис. 2.11). В покое толстые миозиновые нити лишь на небольшое расстояние входят в промежутки между актиновыми. Поэтому в средней части А-диска имеется более светлая Н-зона, где актиновые нити отсутствуют. При электронной микроскопии в ее центре видна очень тонкая М-линия. Она образована цепями опорных белков, к которым крепятся миозиновые протофибриллы. Толщина мышечных волокон зависит главным образом от количества и поперечного сечения миофибрилл.
Механизм мышечного сокращения
При световой микроскопии было замечено, что в момент сокращения ширина А-диска не уменьшается, а I-диски и Н-зоны саркомеров суживаются. Ряд фактов был установлен с помощью электронной микроскопии. Наиболее важно то, что длина нитей актина и миозина в момент сокращения не изменяется. Поэтому Хаксли и Хэнсон разработали теорию скольжения нитей.
Согласно этой теории мышца укорачивается в результате движения тонких актиновых нитей в промежутки между миозиновыми.
Это приводит к укорочению каждого саркомера, образующего миофибриллы. Скольжение же нитей обусловлено тем, что при переходе в активное состояние головки отростков миозина связываются с центрами актиновых нитей и вызывают их движение относительно себя ( рис. 2.12).

Рис. 2.12. Скольжение актиновых и миозиновых нитей в процессе
мышечного сокращения
Сокращение начинается с того, что в области концевой пластинки двигательного нерва возникает потенциал действия (ПД). Он с большой скоростью распространяется по сарколемме и переходит с нее по системе поперечных трубочек саркоплазматического ретикулума (СР), на продольные трубочки и цистерны. Возникает деполяризация мембраны цистерн и из них в саркоплазму высвобождаются ионы кальция (Са2+). На нитях актина расположены молекулы еще двух белков – тропонина и тропомиозина. В состоянии покоя при низкой (менее 10-8 М) концентрации Са2+, тропомиозин блокирует присоединение мостиков миозина к нитям актина. Когда ионы Са2+ начинают выходить из СР (цистерн), молекула тропонина изменяет свою форму таким образом, что освобождаются головки миозина и начинается скольжение за счет ритмического замыкания и размыкания поперечных мостиков миозина с нитями актина. При этом головки ритмически продвигаются по нитям актина к
Z- мембранам.
Передача сигнала от возбужденной мембраны к миофибриллам называется электромеханическим сопряжением. Когда генерация ПД прекращается и мембранный потенциал возвращается к исходному уровню, начинает работать Са2+-насос (фермент Са-АТФаза). Ионы Са2+ вновь закачиваются в цистерны саркоплазматического ретикулума и их концентрация падает ниже 10-8 М. Молекулы тропонина приобретают исходную форму, и тропомиозин вновь начинает блокировать активные центры актина. Головки миозина отсоединяются от активных центров актина, и мышца за счет эластичности приходит в исходное расслабленное состояние.

Рис. 2.13. Механизм сопряжения возбуждения и сокращения [5]
1 – поперечная трубочка саркоплазматической мембраны, 2 – саркоплазматический
ретикулум, 3 – ион Са2+, 4 – молекула тропонина, 5 – молекула тропомиозина
Существенную роль в сократительной активности мышц играют ионы Ca2+. Они активны в саркоплазме при очень низкой (10-6М и менее) концентрации (даже в воде двойной дистилляции концентрация Ca2+ выше, чем 10-6 М). Скелетная мышца не сокращается в ответ на деполяризацию мембраны, если исчерпаны запасы кальция во внутренних депо, а подвергнутые предварительной экстракции препараты волокон скелетной мышцы не сокращаются при добавлении АТФ, если отсутствует Са2+).
Достаточно часто сравнивают процесс сократительного акта в саркомере с движениями гребцов в академической лодке. От филаментов миозина с обеих сторон, подобно веслам в лодке, отходят отростки – миозиновые мостики. В результате реакции на нервный сигнал эти мостики временно пристыковываются к филаментам актина (замыкание), а затем отводятся в позицию под углом 450. За счет этих движений филаменты актина перемещаются между филаментами миозина. После выполнения «гребка» отростки миозина, отрываются от актина и возвращаются в исходное положение (размыкание). За один цикл «замыкания – размыкания» саркомер укорачивается примерно на 1%своей длины. Следовательно, для эффективного напряжения мышцы требуется большое число таких циклов соединения филаментов. В зависимости от типа мышечного волокна и требуемой величины напряжения мотонейроны генерируют импульсы с частотой от 7 до более чем 50 импульсов в секунду.
Саркомеры, расположенных по ходу миофибриллы, включаются последовательно, в процессе работы мышцы их единичные минимальные сокращения суммируются, и миофибрилла сокращается на 25–30%. За счет плотной упаковки миофибрилл, их относительно небольшие сократительные силы складываются в суммарную силу мышечного волокна и в итоге в мышцы.
При непосредственном раздражении мышцы (прямое раздражение) или опосредовано через иннервирующий ее двигательный нерв (непрямое раздражение) одиночным стимулом возникает одиночное мышечное сокращение (рис. 2.14), в котором выделяют три фазы:
- латентный период – время от начала действия раздражителя до начала ответной реакции;
- фаза сокращения (фаза укорочения);
- фаза расслабления.
В режиме одиночного сокращения мышца способна работать длительное время без утомления, но его сила незначительна. Поэтому в организме такие сокращения встречаются редко, например, так могут сокращаться быстрые глазодвигательные мышцы. Чаще всего одиночные сокращения суммируются.
Суммация – это сложение двух последовательных сокращений мышцы при нанесении на нее двух пороговых или сверхпороговых раздражений.
Различают 2 вида суммации: полную и неполнуюсуммацию.
Неполная суммация возникает в том случае, если повторное раздражение наносится на мышцу в начало фазы расслабления.
Полная суммация возникает тогда, когда повторное раздражение действует на мышцу до начала периода расслабления, т. е. в конце периода укорочения.
Амплитуда сокращения при полной суммации выше, чем неполной.

Рис. 2.14. Одиночное и тетаническое сокращение мышцы [6]
В естественных условиях к скелетной мышце из ЦНС поступают не одиночные импульсы, а серия импульсов, следующих друг за другом с определенными интервалами, на которую мышца отвечает длительным сокращением. Такое длительное сокращение мышцы, возникающее в ответ на ритмическое раздражение, получило название тетанического сокращения или тетануса.
Различают два вида тетануса: зубчатый и гладкий.
Если каждый последующий импульс возбуждения поступает к мышце в тот период, когда она находится в фазе укорочения, то возникает гладкий тетанус, а если в фазу расслабления – зубчатый тетанус (рис. 2.14).
Амплитуда тетанического сокращения превышает амплитуду одиночного мышечного сокращения. Исходя из этого, Гельмгольц объяснил процесс тетанического сокращения простой суперпозицией, т. е. простой суммацией амплитуды одного мышечного сокращения с амплитудой другого. Однако в дальнейшем было показано, что при тетанусе имеет место не простое сложение двух механических эффектов, т. к. эта сумма может быть то большей, то меньшей. Н. Е. Введенский объяснил это явление с точки зрения состояния возбудимости мышцы, введя понятие об оптимуме и пессимуме частоты раздражения.
Оптимальной называется такая частота раздражения, при которой каждое последующее раздражение осуществляется в фазу повышенной возбудимости. Тетанус при этом будет максимальным по амплитуде – оптимальным.
Пессимальной называется такая частота раздражения, при которой каждое последующее раздражение осуществляется в фазу пониженной возбудимости. Тетанус при этом будет минимальным по амплитуде – пессимальным
Режимы мышечных сокращений. Различают изотонический, изометрический и смешанный режимы сокращения мышц.
При изотоническом сокращении мышцы происходит изменение ее длины, а напряжение остается постоянным. Такое сокращение происходит в том случае, когда мышца не перемещает груз. В естественных условиях близкими к изотоническому типу сокращений являются сокращения мышц языка.
При изометрическом сокращении длина мышечных волокон остается постоянной, меняется напряжение мышцы. Такое сокращение мышцы можно получить при попытке поднять непосильный груз.
В целом организме сокращения мышц никогда не бывают чисто изотоническим или изометрическим, они всегда имеют смешанный характер, т. е. происходит изменение и длины, и напряжения мышцы. Такой режим сокращения называется ауксотоническим, если преобладает напряжение мышцы, или ауксометрическим, если преобладает укорочение.
Зависимость величины работы от нагрузки подчиняется закону средних нагрузок. При увеличении нагрузки работа мышц первоначально растет. При средних нагрузках работа становится максимальной. Если увеличение нагрузки продолжается, то работа снижается.
Такое же влияние на величину работы оказывает ее ритм. Максимальная работа мышцы осуществляется при среднем ритме.
Вопросы для самоконтроля
1. Какие вы знаете физиологические свойства скелетных мышц?
2. Что такое двигательная единица?
3. Какие существуют виды двигательных единиц? В чем их особенности?
4. Что такое композиция мышц? В чем ее значимость для практики спорта?
5. От чего зависит сила мышцы?
6. В чем суть внутримышечной и межмышечной координации?
7. Чем различаются одиночное и тетаническое сокращение?
8. Какие существуют режимы мышечного сокращения?
Глава 3. Центральная нервная система
3.1. Функции ЦНС. Рефлекторный механизм
деятельности ЦНС
Организм человека представляет собой совокупность разных структур, отличающихся по строению, роли и степени участия в процессах жизнедеятельности. Выживание в окружающей среде требует согласованной работы этих структур – тканей, органов и совокупностей органов. Такое согласование осуществляют интегрирующие системы – система желез внутренней секреции, иммунная и нервная системы. В настоящее время считают, что центральное место в этой триаде занимает нервная система. Это определяется целым рядом обстоятельств, таких, как возможность точного воздействия на определенные структуры, высокая скорость выполнения регулирующих воздействий, очень точная градация силы влияний и ряд других особенностей регуляции с помощью нервной системы.
Нервная система человека включает в себя центральную нервную систему (ЦНС) и периферическую нервную систему (ПНС). ЦНС включает головной и спинной мозг, а ПНС, обеспечивающая связь ЦНС с различными частями тела, – черепно-мозговые и спинномозговые нервы, а также нервные узлы (ганглии) и нервные сплетения, лежащие вне спинного и головного мозга.
Центральная нервная система (ЦНС) выполняет функцию интеграции всех структур организма в единое целое. Она регулирует все процессы, происходящие в организме, обеспечивая индивидуальное приспособление его к изменяющимся условиям существования: Основные функции ЦНС:
1) воспринимает действие на организм разнообразных раздражителей;
2) производит анализ и синтез раздражений, а затем формирует поток центробежных нервных импульсов, под влиянием которых изменяется работа тех или иных органов;
3) определяет поведение человека и животного, его взаимоотношения с внешней средой.
Значительную сложность представляет исследование ЦНС, ведь исследуется не просто сверхсложная система, объект исследования выполняет и роль исследовательского инструмента.
Основными методами изучения функций ЦНС являются:
- удаления и раздражения (в клинике и на животных);
- регистрация электрических явлений (электроэнцефалограмма – регистрация суммарной электрической активности различных областей мозга);
- метод условных рефлексов;
- компьютерная томография (морфофункциональные изменения мозга на различной его глубине) и многие другие.
Нервная система состоит из трех основных клеточных компонентов:
1) нервных клеток (нейронов);
2) связанных с ними клеток нейроглии;
3) клеток соединительной ткани.
Глиальные клетки составляют специфическое микроокружение для нейронов, обеспечивая условия для генерации и передачи нервных импульсов, а также осуществляя часть метаболических процессов самого нейрона. Нейроглия выполняет опорную, трофическую, секреторную, разграничительную и защитную функции.
Нейрон является морфофункциональной единицей центральной нервной системы (ЦНС) (рис. 3.1). Деятельность нейрона заключается в выработке нервных импульсов. Проведение возбуждения осуществляется электрическим путем.
 |
Рис 3.1. Схема строения нейрона
Каждая нервная клетка состоит из тела, множества коротких отростков – дендритов и чаще всего одного длинного отростка – аксона. Тело и дендриты играют основную роль в восприятии и переработке сигналов поступающих от других нейронов. Тело клетки выполняет питательную функцию. Функции аксона состоит в проведении нервных импульсов к другим нейронам или к исполнительным органам – мышцам, железам и т. п.
Передача возбуждения с одного нейрона на другой осуществляется при помощи синапса (рис. 3.2). Аксоны могут образовывать синапсы на теле другого нейрона или на его отростках.



|
|
|
|
|
|
|

Рис. 3.2. Строение синапса [7]
В наиболее общем (и весьма упрощенном виде) синапс имеет вид бляшки. Он состоит из пресинаптической мембраны, синаптической щели и постсинаптической мембраны. В зависимости от механизма передачи нервного импульса различают химические и электрические синапсы.
В химическом синапсе внутри бляшек находятся пузырьки с медиатором (нейротрансмиттером ). Медиаторы – это биологически активные химические вещества, с помощью которых осуществляется передача электрического импульса с нервной клетки через синаптическое пространство между нейронами. По химической природе это аминокислоты, пептиды, моноамины (в том числе катехоламины). В настоящее время известно более 30 разновидностей медиаторов.
При достижении нервным импульсом пресинаптической мембраны происходит высвобождение медиатора в синаптическую щель. Диффундируя, медиатор попадает на постсинаптическую мембрану и, взаимодействуя с ней, изменяет проницаемость постсинаптической мембраны для ионов Na+ и K+, как это описано выше, в результате чего развивается деполяризации и возникает потенциал действия, распространяющийся по нейрону и его отросткам.
В электрическом синапсе клетки соединяются высокопроницаемыми контактами с помощью особых белковых субъединиц (коннексонов), через которые передается электрический сигнал.
По функциональному признаку можно выделить три группы нейронов:
1) чувствительные (сенсорные или афферентные), воспринимающие своими окончаниями раздражение и передающие его в ЦНС. Эти нейроны расположены вне ЦНС, в спинномозговых ганглиях или в аналогичных ганглиях черепно-мозговых нервов;
2) промежуточные или вставочные нейроны – осуществляют контакты между нервными клетками;
3) эффекторные или эфферентные нейроны – обеспечивают эффект деятельности нервной системы, посылая импульсы к рабочим органам.
В ЦНС существуют возбуждающие (работа описана выше) и тормозящие, или тормозные нейроны. Отличие этих нейронов состоит в том, что медиатор, выделяющийся из пресинаптических мембран, приводит к развитию гиперполяризации и препятствует распространению возбуждения.
По классическим физиологическим представлениям основным механизмом деятельности ЦНС является рефлекторный. Нервная система под влиянием внешних или внутренних воздействий (стимулов) приходит в состояние возбуждения, из ЦНС это возбуждение передается определенному органу, который отвечает на это возбуждение изменением своих функций. Результатом рефлекторной деятельности могут являться начало, усиление или, наоборот, ослабление какой-либо деятельности организма. Изменение функционального состояния органов и систем также происходит по механизму рефлекса.
Рефлекс (отражение) – это стереотипная закономерная реакция организма на раздражение, осуществляемая при непосредственном участии ЦНС в ответ на раздражение рецепторов.
Впервые отражающий (рефлекторный) принцип деятельности нервной системы был предложен французским естествоиспытателем и философом
Р. Декартом («Рассуждение о методе», «Страсти души», 1637). Он предложил схему рефлекса для объяснения так называемых непроизвольных движений. Термин рефлекс был впервые предложен чешским естествоиспытателем
И. Прохаска (1749–1820). В XIX веке Ч. Белл и Ф. Мажанди показали наличие анатомической основы рефлекса. М. Холл предложил термин «рефлекторная дуга». По мнению всех этих исследователей, рефлекторный механизм свойственен только спинному мозгу.
Эти взгляды были пересмотрены великим русским физиологом
И. М. Сеченовым (1829–1905), распространившим рефлекторный принцип на работу всей нервной системы, включая головной мозг. И. М. Сеченов сформулировал главные положения рефлекторной теории.
Крупнейшие достижения в изучении рефлекторной деятельности принадлежат русскому физиологу И. П. Павлову. Теоретическим обоснованием рефлекторной теории являются выделенные И. П. Павловымосновные принципы рефлекторной теории:
- принцип детерминизма – каждый процесс в головном мозге вызывается изменениями, происходящими вне или внутри организма;
- принцип анализа и синтеза – заключается в том, что кора, благодаря анализу, способна различать, дифференцировать раздражения, выделяя из них те, на которые следует дать реакцию в настоящий момент, а благодаря синтезу, способна объединять, синтезировать эти выделенные раздражения в ответные реакции;
- принцип структурности означает, что функции головного мозга связаны с его строением, что всякий нервный процесс происходит в определенных морфологических образованиях.
Классифицировать рефлексы можно на основе разных признаков. Можно их поделить в зависимости от того, в каком отделе мозга они замыкаются. Более правильно все же выделять лишь отдел мозга или нервный центр, обязательно участвующий в осуществлении данного рефлекса.
Работы И. П. Павлова позволили выделить безусловные и условные рефлексы.
Структурной основой рефлекса является рефлекторная дуга (рис. 3.3.). Для осуществления рефлекса необходимо наличие всех звеньеврефлекторной дуги: сенсорных рецепторов; афферентных или чувствительных нервных проводников; нервных центров; эфферентных или двигательных нервных проводников; эффекторов или исполнительных органов.
В простейшем случае рефлекторная дуга может состоять всего из двух нейронов.
 | |||||||||||||

Рис. 3.3. Схема рефлекторной дуги [8]
( 1 – чувствительный (афферентный) нейрон; 2 – вставочный (кондукторный) нейрон;
3 – двигательный (эфферентный) нейрон; 4 – нервные волокна тонкого и клиновидного пучков; 5 – волокна корково-спинномозгового пути).
Рецепторы – сложные образования (состоят из терминалей дендритов, чувствительных нейронов, включая клетки глии, образований межклеточного вещества и специализированных клеток других тканей), которые обеспечивают превращение влияния стимулов (раздражителей) внешней или внутренней среды в нервный импульс. В некоторых рецепторах (вкусовых и слуховых рецепторах человека) раздражитель непосредственно воспринимается специализированными клетками.
Область, где расположены рецепторы, при раздражении которых возникает данный рефлекс, называется рефлексогенной зоной, или рецептивным полем. Каждый рефлекс имеет свою рефлексогенную зону.
Типы рецепторов:
- сенсорные рецепторы (восприятие нервной системой различных раздражителей внешней и внутренней среды) (табл. 3.1).
- клеточные химические рецепторы (восприятие информации, переносимой молекулами химических веществ).
Общим свойством рецепторов является способность преобразовывать один вид энергии в другой, т. е. в энергию биопотенциала.
Таблица 3.1
Сенсорные рецепторы
| По характеру восприятия | По источнику воспринимаемой информации |
| Механорецепторы Хеморецепторы Терморецепторы Ноцицепторы | Экстерорецепторы (информация из внешней среды) Интерорецепторы (из внутренней среды) |
Важнейшим звеном рефлекторной дуги являются – афферентные и эфферентные нервные проводники.
Основной функцией нервов является передача сигналов к нервному центру от рецепторов (афферентные) или от нервного центра к эффектору (эфферентные).
Собственно проводниками являются нервные волокна, входящие в состав периферических нервов или белого вещества головного и спинного мозга.
Нервные волокна различаются:
1) толщиной (диаметром);
2) наличием или отсутствием миелиновой оболочки;
3) скоростью проведения возбуждения;
4) длительностью потенциалов действия;
5) продолжительностью следовых потенциалов.
Механизм проведения и возбуждения в нервных волокнах объясняется возникновением локальных токов, появляющихся между возбужденными и невозбужденными участками мембраны нервного волокна.
При этом в безмиелиновых волокнах возбуждение распространяется непрерывно, а в миелинизированных волокнах – скачками, между перехватами Ранвье, лишенными миелиновой оболочки (рис. 3.4).
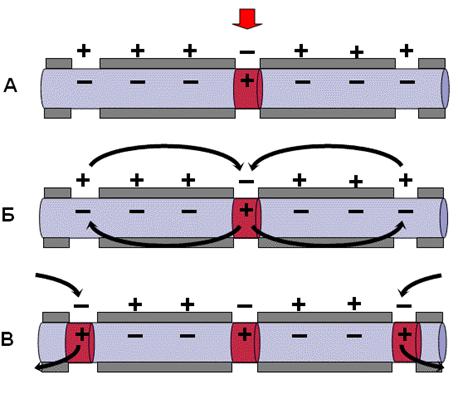
Рис. 3.4. Механизм распространения возбуждения по миелиновому
нервному волокну [9]
3.2. Понятие о нервном центре. Свойства нервных центров
Нервным центром называется функциональная совокупность нейронов, необходимая для осуществления определенного рефлекса или регуляции той или иной функции.