Да и первый нередко встает из-за стола, чтобы вновь вернуться к нему, найдя удачный оборот речи или на ходу додумав мысль. Варианты, зависящие от индивидуальных свойств, настроения, воли, привычки человека. Но как многое от этого не зависит. Трудные или легкие, жизненно важные или почти незначимые вопросы задает человеку жизнь в меняющихся условиях внешней среды, при разных состояниях организма и мозга. Мозг его должен решать их - хуже или лучше - ив тишине кабинета, и в шуме толпы. Гибкость мозга, его способность приспосабливаться к условиям, и обеспечивает ему огромный аппарат гибких звеньев системы.
Эти почти натурфилософские рассуждения могли бы быть поводом к научным исследованиям, могли сами по себе лечь в основу предположения о возможной сложности принципов организации мозговых систем обеспечения психических функций. В данном случае они являются не предпосылкой, а следствием целенаправленного изучения динамики физиологических показателей в различных зонах мозга при психологических пробах, реализуемых в различных условиях внешней среды и при разных внутренних условиях функционирования мозга. Вопрос этот подробно освещался в наших предыдущих статьях и монографиях (Бехтерева, 1971, 1974, 1977). Здесь целесообразно подчеркнуть: хотя гибкие звенья мозговой системы обеспечения психической деятельности в целом и обнаружили динамику в прямой зависимости от условий ее реализации, нельзя утверждать, что, скажем, выполнение теста на краткосрочную вербальную память на фоне мелькающего света обязательно включит строго определенную зону мозга. Оно включит именно ее с известной вероятностью. Так же обстоит дело с зонами мозга, включающимися или выключающимися на фоне других помех: музыки, постороннего разговора и т. п. Иными словами, можно было бы говорить не о мозговой системе обеспечения психической деятельности даже применительно к одному какому-то ее виду, а об организации системы, включающей жесткие элементы разной значимости, симметрично и асимметрично организованные, о своего рода субсистемах гибких звеньев, где дублирование и, таким образом, взаимозаменяемость представлены значительно шире. И наконец, вероятен, хотя еще и не доказан иерархический принцип организации системы и соответственно наличие в ней пейсмекерных механизмов.
Коротко о пейсмекерном механизме. Как, на основе каких механизмов мозга
осуществляется скоростное, а нередко практически одномоментное задействование множества звеньев системы обеспечения психической деятельности? Как можно понять включение в деятельность огромного множества звеньев системы с учетом синаптической задержки (Meyers, 1974)? Каковароль «тесных соединений» нейронов, прямой электротонической передачи (Костюк, 1973; Adey, 1977), электрических синапсов с отсутствием в них синаптической задержки и принципиальной возможностью двусторонней передачи (Воронцов, 19616; Шаповалов, 1975)? Какую роль в нервной системе играет великое множество видов синапсов с самыми разными типами биохимической медиации (Глебов, Крыжановский, 1978)? Каковароль слабых электромагнитных полей в передаче сигналов (Frohlich, 1973; Adey, 1977)? Необходимо ли и оправдано ли введение представлений о пейсмекерном механизме в организации нейрофизиологического обеспечения этой деятельности, естественно, не альтернативных по отношению ко всем здесь поставленным вопросам? В результате исследований мозговой структурно-функциональной организации психической деятельности наряду с общими чертами показано ее отличие от других видов деятельности не только по ее сложности, но и но мозговой организации. Речь идет об относительно большей роли гибких звеньев в этой системе по сравнению с мозговыми системами обеспечения других видов деятельности. В звеньях системы обеспечения психической деятельности важно также учесть роль специфической нейродинамики в структуре наряду с фактором специфичности самой структуры и ее функционального состоя ния (Бехтерева, 1977).
Отдавая себе отчет в первостепенной значимости этой нейродинамики, мы представляем себе, что, если без введения концепции о пейсмекере в организации и реализации психической деятельности обойтись трудно, пейсмекер в этом случае должен быть принципиально отличным от жестко структурно-фиксированных образований и иметь вид первичной нейродинамической констелляции структур, возникающей под влиянием внутренних или внешних факторов среды, в том числе и словесной команды.
Конкретный характер работы и организации пейсмекерного механизма может и должен быть различным в зависимости от рода деятельности, а в осуществлении ее играют важную и разную роль все упомянутые выше механизмы, а возможно, и некоторые другие.
Так, в частности, в поставленных жизнью или психологическим тестом задачах на обобщение первоначальная роль пейсмекера - обеспечение условий расширенного поиска, что, повидимому, решается за счет включения активирующих (синхронизирующих?) влияний. Опознание принадлежности двух-трех слов (явлений) к одному смысловому (ассоциативному) полю определяет направленность и ограничение - торможение - избыточного поиска. Если деятельность повторяется, может быть использована активация матрицы памяти того пейсмекера, который уже обеспечивал аналогичную или близкую по типу деятельность. Если деятельность
изменяется, меняется и сам динамический пейсмекер. В наиболее общем виде можно, повидимому, предположить, что его роль сводится, прежде всего, к управлению, организации и реорганизации системы обеспечения психической деятельности с соответствующей активацией ассоциативных полей долгосрочной памяти и детерминированным ситуацией ограничением или расширением этой активации на разных фазах развития психической деятельности. Наличие такого динамического пейсмекера не исключает присутствия сличающего аппарата - акцептора результатов действия П. К. Анохина (1968). Возможно, что акцептор действия является одной из фаз трансформации, развития пейсмекера.
Представление о пейсмекерном механизме, безусловно, еще нуждается в уточнении путем целенаправленных исследований, которые при всей динамичности пейсмекерной организации психической деятельности все же должны пройти фазу хотя бы приближенного и разного в различных случаях привязывания к структуре. Какие в этом плане могут быть предложены рабочие гипотезы?
Учитывая данные нейропсихологии о том, что глубокие отделы лобных долей обеспечивают избирательность памяти (Лурия, 1977) и те же отделы мозга, а по данным других исследователей, и глубокие отделы височных долей (Brazier, 1966, 1967а, 19676) - план текущей деятельности и реализацию этого плана, можно себе представить, что эти области мозга помимо прямых адресатов, связанных с характером раздражителя, как правило, участвуют в констелляции структур, составляющих динамический пейсмекер. Пейсмекерный механизм включает как специфические, так и неспецифические элементы, причем роль модулирующих неспецифических в этом аппарате, вероятно, очень велика и, в частности, как предпосылка к взаимосвязи нервных элементов. Специфические зоны мозга, участвующие в пейсмекерной организации, прежде всего те, куда адресуется раздражитель, явившийся поводом к развитию деятельности. Пейсмекер, как указывалось, детерминирует ассоциативный поиск, исследовательские этапы психической деятельности и соответствующий поиск в долгосрочной памяти.
Что нового дают эти представления? Прежде всего новой является сама гипотеза о возможности возникновения или активации пейсмекера под влиянием событий внутренней и внешней, в том числе и социальной, среды. Новыми являются также представления о его формирующейся ex tempore динамической констелляции структур и, что принципиально, об отсутствии жестко предопределенной (генетически) закрепленности пейсмекера за структурой.
Представление о динамической пейсмекерной организации выгодно для понимания мозгового обеспечения громадного разнообразия психических процессов. Именно динамичность в обеспечении психики, как это ни парадоксально на первый взгляд, определяет не только его гибкость, но и экономичность. При чисто структурных пейсмекерах, генетически запрограммированных и индивидуально развернутых, количество их должно было бы быть астрономическим. Кроме того, сам процесс
индивидуального формирования и развития психической деятельности совместим только с гипотезой и реальностью динамических пейсмекеров. Структурный пейсмекер - допустимый элемент лишь генетически запрограммированной деятельности, развертывающейся, а не развивающейся в течение жизни.
Прогресс как результат творческой психической деятельности человека возможен, если эта деятельность базируется на принципе динамических управляющих аппаратов, количество и организация которых определяются нуждами деятельности. Фазовое развитие и реорганизация динамических пейсмекеров в соответствии с динамикой деятельности определяют логику мыслительных процессов или являются нейрофизиологическим механизмом этой логики, предотвращая недетерминированный хаос ассоциаций. Передача закодированного возбуждения от пейсмекерной системы идет далее к другим звеньям психической деятельности, включая первоначально наиболее значимые нейронные популяции или распространяясь сразу на все зоны мозга, имеющие отношение к психической деятельности. В этой передаче несомненно важную роль играет синаптический аппарат. Однако даже исключительное богатство внутримозговых связей не может, повидимому, обеспечить достаточно быстрое и, что особенно важно, одновременное включение всех необходимых для сложной психической деятельности звеньев мозговой системы ее обеспечения. Скоростное включение обычно первоначально избыточного числа мозговых звеньев укладывается в высказанные нами в 1971 году представления о возможности одномоментной реализации только одного вида сложной психической деятельности.
Эти представления, равно как и признание необходимости скоростной передачи возбуждения на звенья системы обеспечения психической деятельности, ставят альтернативу: или пейсмекерный аппарат оказывается в состоянии передать одновременно обычным синаптическим путем сигналы всем звеньям системы обеспечения психической деятельности в связи с созданием условий идеального распространения возбуждения, или в мозгу вообще может существовать и другая, более адекватная по временному фактору форма одновременной взаимосвязанной активации многих элементов, особенно важная для описанной ситуации.
В пользу первого положения могут свидетельствовать результаты исследований и теоретические представления М. Н. Ливанова (1975) о синхронизирующей роли неспецифических влияний в головном мозгу, обеспечивающих оптимизацию условий внутримозгового взаимодействия. Другие гипотезы, по-видимому, также стоит исследовать, используя в качестве опорных теоретических и фактических оснований представления Богоша (Bogoch, 1968, 1973), И. С. Бериташвили (1969), В. С. Русинова (1969), Н. В. Голикова (1970), Эди (Adey, 1977), атакже некоторые данные нашей лаборатории (Илюхина, 1977). Синаптические приборы следует рассматривать шире, чем только как передающий аппарат. Они - и аппарат селекции и, вероятно, квантующий аппарат.
Безусловно, необходимо шире изучать пути и принципы передачи сигналов в мозгу. На сегодня в качестве рабочих представлений мы принимаем, что пространственновременные взаимоотношения осуществляются в мозгу в процессе реализации психической деятельности за счет последовательности и, таким образом, взаимодействий синаптической (задержанной) и других форм передачи импульсов, немалую роль в которых могут играть и сверхслабые градуальные процессы.
Доминирование пространственного фактора (одновременное задействование большого числа звеньев), полученное за счет скоростной передачи, является не потерей фактора времени, а целесообразным уменьшением задержек, своего рода переходом на другой порядок отсчета времени, соизмеримый в биологических системах со временем прохождения импульсов по нервному проводнику. На уровне популяций нейронов фактор времени вновь особенно значимо проявляется в квантованных интервалах групповых последовательностей импульсов, отражающих перестройки пространственных отношений. При этом фактор пространства проявляется во взаимодействии отдельных близлежащих нейронов (микроуровень) и звеньев системы (макроуровень) (Бехтерева и др., 1977а).
Вопрос об удельном значении различных форм передачи импульсов в нервной системе дискуссионный. Однако фокусирование внимания исследователей на них неизбежно при попытках понять деятельность целого мозга и необходимо для преодоления разрыва между двумя основными современными направлениями изучения различных биологических объектов и, в частности, нервной системы - исследованием свойств отдельной нервной клетки и сложнейшей деятельности всего мозга человека.
Все это важно, и все же, занимаясь физиологией психической деятельности человека со всеми ее особенностями быстродействия, не зависящего от пространства, невольно начинаешь думать о том, нет ли в мозгу механизма, аналогичного понятию когерентной работы, некой системы элементов, как в физике твердого тела. Я, кажется, не одинока с этой, ставшей почти навязчивой идеей. Именно об этом, повидимому, думает и Эди, упоминая в статьях о мозге о «туннельном эффекте», таком характерном для полупроводниковых диодов (Adey, 1977). Или невольно за этими словами видится больше, чем в них содержится?
Несомненно, можно и нужно продолжать выявление в мозгу зон, участвующих в обеспечении психики, изучать структурно-функциональную организацию мозга. Важно накапливать и анализировать все больше сведений об участии различных мозговых структур в обеспечении разных проявлений и аспектов психической деятельности. Такого рода данные получают у нас в отделе (Смирнов, 1976; Шандурина, Калягина, 1979), они появляются в литературе (Walter et al., 1978). Но принципиальный, опорный ответ на вопрос, каким образом организовано в мозгу структурно-функциональное обеспечение психической деятельности, по-видимому, уже дан (Бехтерева, 1974).
В конце 60-х годов в качестве важнейшей задачи в изучении мозгового обеспечения психической деятельности была определена расшифровка протекающих в корковых и подкорковых структурах процессов в момент, когда человек слышит и опознает хотя бы простейшее слово. Такая задача представлялась на том уровне исследования физиологии мозга человека сложной до неразрешимости. Между нею и тем, что было уже сделано, стоял хотя и невидимый, но вполне ощутимый не только технический, но и психологический барьер, обусловленный прежде всего невозможностью решить эту задачу традиционными математико-техническими приемами, использовавшимися в тот период в биологии и медицине. Своего рода комплексная идейная неподготовленность исследователей. Эта неподготовленность определялась и отставшей от фактов предубежденностью о правомерности поисков тонких коррелятов психических явлений только в коре, естественно, без указания точного адреса поисков, малочисленностью исследовательских групп, работающих в области физиологии человека с использованием комплекса адекватных физиологических методик.
Основой изучения этого вопроса в нашей лаборатории был многолетний опыт исследования мозга с помощью комплексного метода и также многолетняя (почти с «доисторических» времен - с 1954 года!) совместная работа физиологов и врачей со специалистами технического профиля.
Импульсная активность нейронных популяций при мыслительной деятельности
Первый этап исследования. Мы имели возможность находить точки мозга, более или менее тесно связанные с психической деятельностью. Для расшифровки протекающих в них процессов физиологу предстояла сначала задача выбрать из регистрируемых биоэлектрических процессов - наиболее подходящий для этой цели. Первым (хотя и не единственным) кандидатом на данную роль была импульсная активность нейронов, прежде всего, в связи с быстродействием мыслительных процессов. Далее встал вопрос: отдельная нервная клетка или функционально объединенное сообщество нервных клеток? Многолетний опыт экспериментальных исследований (Василевский, 1968; Альтман, 1972, и др.) показал, что одна нервная клетка чаще всего не способна отражать своей активностью сложные сигналы. При регистрации динамики отдельной или, точнее, отдельных клеток как представительных единиц в человеческом мозгу можно получить ряд интересных сведений о его работе (см. выше). Однако на уровне группы нервных клеток, работающих в том нужном согласии друг с другом, которое создает новое качество, можно было надеяться найти отражение уже более сложных сигналов (Коган, 1973). Вопросы о свойствах популяции нервных клеток подробно рассматриваются Буллоком (Bullok, 1979).
Таким образом, еще в период, когда сравнительно редко использовалось отведение импульсной активности не с микроэлектродов, а с так называемых микромакроэлектродов (в нашем случае с диаметром 50-100 мкм и рабочей поверхностью от 0.01 до 0.15 мм), было начато изучение реорганизации импульсной активности нейронных ансамблей при простейших видах мыслительной деятельности. С вживленных в различные структуры мозга (кору, образования таламуса и стриопаллидарной системы, верхние отделы ствола, медиобазальные отделы височных долей) электродов регистрировалась активность популяций нервных клеток. В этой активности обычно обнаруживались пики разрядов нескольких нервных клеток, более или менее существенно превышавшие по амплитуде труднодифференцируемую активность большого количества нервных клеток. Варианты регистрируемой с вживленных электродов активности представлены на рис. 14.
Любое исследование тонких нейрофизиологических коррелятов мыслительной деятельности до последнего времени имело и еще, по-видимому, в течение ряда лет будет иметь, кроме собственно нейрофизиологических, по крайней мере два аспекта. Так, во-первых, в каждом из них в соответствии с нейрофизиологическими задачами требовалось найти адекватный психологический подход. И хотя многие из применявшихся тестов строились на основе классических психологических проб, далеко не все могло быть использовано в традиционном варианте (тест Вине и др.). Во-вторых, возможности нейрофизиологического анализа ответов активной, реагирующей на психологический тест точки существенно зависели от степени совершенства математико-технической обработки извлечения полезного сигнала.
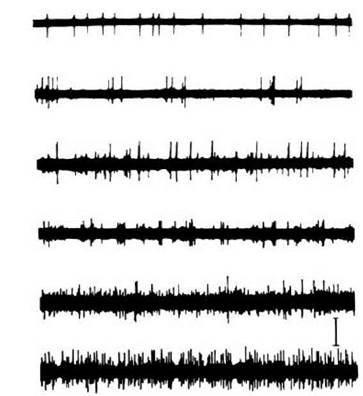
Рис. 14. Примеры
нейронограммы мультиклеточной активности различных зон коры и подкорковых структур головного мозга человека.
Сверху вниз:
поясная извилина, премоторная область коры, вентролатеральное таламическое ядро срединный центр, заднее вентральное латеральное таламическое ядро, бледный шар. Калибровка: 20 мс, 50 мкВ
Для избежания разночтений уточним, в чем разница между популяцией нервных клеток и их рабочим ансамблем. С электрода регистрируется активность группы нервных клеток. Это - всегда популяция, но не всегда ансамбль. Как можно проверить подобное утверждение? Дается задача, одна, другая - сосчитать что-то в уме, повторить ряд слов. Затем с помощью специальных программ и ЭВМ изучается реакция этой популяции нервных клеток. Какие здесь возможны варианты? Может просто не произойти ничего существенного - и тогда, скорее всего, данная популяция не имеет отношения к реализуемой деятельности. Реакция может быть относительно простой - в виде общего тонического увеличения или уменьшения частоты разрядов.
В этом случае следует предположить, что данная популяция вносит в систему обеспечения психической деятельности «энергетический» вклад того или другого знака или что существующие методы еще не способны обнаружить более тонкие изменения. Может выявиться, наоборот, перестройка организации работы нервных клеток. И тогда правомерно использовать термин «ансамбль», хотя в конкретном
случае это может быть лишь часть ансамбля, так как остальная находится за пределами возможностей регистрации активности ее клеток с данного электрода. А может быть и иначе. Только часть популяции существенно реорганизуется, работает в данных условиях как слаженный ансамбль. Но вот задается другая деятельность, ансамбль захватывает другие клетки из популяции или, наоборот, становится меньше. Ансамбли очень динамичны, они все время «дышат». Наверное, тем, кто не работает непосредственно в данной области, представить себе организацию и реорганизацию ансамблей можно, вспомнив динамичную световую рекламу, - из набора однотипных источников света слагаются ежемоментно меняющиеся, иногда очень сложные световые узоры. Понятно, что не только это сравнение, но и приведенные выше положения - лишь схема, которую иногда очень нелегко уточнить в процессе математико-технической обработки.
Те зоны, в которых нами обнаруживались воспроизводимые изменения физиологических показателей мозга при психологических пробах, не относились к анатомическим образованиям акустического (слухового) анализатора. Лишь теперь, через много лет после начала наших исследований, у тех, кто работает в области мозгового анализа звуковых сигналов, начал пробуждаться интерес именно к этим зонам, в которых, как предполагают Эммере (Emmers, 1970) и Я. А. Альтман с соавторами (1979), происходит обработка сложных сигналов, высшая по отношению к протекающей в классических образованиях анализатора. И, как считает Б. Ф.
Толкунов (1978), именно в этих зонах существуют оптимальные анатомофизиологические условия для высшей интеграции. Тем не менее самым началом нашего углубленного изучения тонких нейрофизиологических коррелятов психической деятельности были поиск и отражения акустических характеристик слов и их элементов в активности популяций (ансамблей) нейронов структур стриопаллидарной системы, таламуса, медиобазальных отделов височных долей, верхних отделов ретикулярной формации не как дань представлениям Пенфильда (Penfield, 1975) о центроэнцефальной системе, а как результат собственного опыта. Как указывалось, именно в этих зонах отмечались воспроизводимые реакции при психологических пробах. При этом мы отдавали себе отчет в том, что поиск коррелятов может быть затруднен непрямым отражением акустических свойств сигнала в связи с характером образований.
В результате первых исследований, по ходу которых проводилась весьма трудоемкая отработка методов анализа мультиклеточной активности, была показана, хотя и непрямая, связь развивающихся в области этих подкорковых структур пространственно-временных перестроек импульсной ак тивности с частотноамплитудными характеристиками входного сложного звукового сигнала - слова (Бехтерева и др., 1971). Известную роль в прогрессе исследований сыграло предложенное на основе работ Сондерсона с соавторами (Sanderson et al., 1973) и П.
В. Бундзена (1976) использование так называемой дистрибутивной формы
представления пространственно-временны^х перестроек импульсной активности нейронных популяций. Эта форма не только открыла новые возможности машинного анализа импульсной активности при психологических пробах, но и явилась необходимым методическим толчком для изучения значимых элементов в перестройках мультиклеточной активности. Детали математико-технической обработки импульсной активности приведены в работах П. В. Бундзена с соавторами (19756), Ю. Л. Гоголицына, П. Д. Перепелкина (1975), Ю. Д. Кропотова (1976, 1978), П. Д. Перепелкина (1977, 1978), в разделе П. Д. Перепелкина в работе Н. П.
Бехтеревой с соавторами (1977а). Идет постоянное совершенствование методов на основе общей принципиальной схемы анализа. По ходу изложения материалов там, где без этого нельзя обойтись, будет приведена физиологическая логика этого математико-технического анализа.
Как видно из наших первых работ, слово как сложный акустический сигнал и - что в этих работах не исключается, но и не доказывается, - как сигнал семантически значимый вызывает тонические и фазические перестройки импульсной активности. Одновременно в данных работах показано, что упомянутые перестройки обнаруживаются только в части популяций, а ансамблем может являться часть большой популяции. Как это обнаруживается? Если при работе с отдельными нервными клетками для получения представления о зоне мозга исследователи идут неизбежно по пути накопления данных о поведении нейронов в этой зоне, то при работе с популяциями нервных клеток методической основой последующей интегративной фазы, объединения, является первоначальное разделение популяций на достаточно малые группы нейронов. Осуществляется выделение тех разрядов нейронов, которые, превышая уровень физиологического шума, позволяли проводить исследования при минимально возможных искажениях мозговых сигналов - имеется в виду прежде всего искажение формы разряда при регистрации в результате суперпозиции двух и более спайков. С этой целью мы прежде всего разделяли разряды по амплитуде. Такой метод при некоторой его уязвимости и используется не только у нас, ной в ряде других исследований (Keidel, 1975; Schwartz et al., 1976; Halgren et al., 1978; Ravagnati et al., 1979). Разделение по амплитуде производится в виде так называемой «оконной» дискриминации (Бехтерева и др., 1973), предполагающей выделение нескольких амплитудных уровней разрядов. При этом в каждое «окно» попадают именно те разряды, пик которых оканчивается в его пределах. Именно на основе этой оконной дискриминации и были разделены разряды мультиклеточной активности в наших первых исследованиях.
Наравне с «оконной» дискриминацией в связи с накоплением достаточного материала и отсюда возможности выбора сейчас предпочтительно используется определенный амплитудный порог, зависящий от соотношения амплитуд выделяемых отдельных разрядов и физиологического шума более низкой амплитуды, «щетки»,
искажающих друг друга разрядов многих клеток; обычно этот порог определяется в размере 0.7 от максимальной амплитуды разрядов. При этом, естественно, учитывается лишь часть регистрируемой активности, но, что очень важно, ее наименее искаженная или даже практически неискаженная часть.
В обоих случаях обычно оказывается возможным обнаружить большее или меньшее количество разрядов равной и разной амплитуды. При исследовании активности отдельных нейронов амплитудный показатель может до известной степени и на небольшом промежутке времени помочь идентифицировать, опознать нейрон. Сходно обстоит дело и при дифференциации мультиклеточной активности. Однако идентификация отдельных нейронов более надежна, хотя тоже не абсолютна, при использовании и амплитудного, и дополнительного показателя - формы потенциалов действия. Этот прием также с 1973 года в течение ряда лет использовался в нашей лаборатории.
При регистрации мультиклеточной активности оказывается возможным выделить с небольшими искажениями динамику активности до четырех, редко более, отдельных нейронов, то есть принципиально то же, что возможно и в условиях аналогичного собственно микроэлектродного исследования.
Существенный выигрыш за счет регистрации именно мультиклеточной активности, а не активности отдельных нейронов, связан с возможностью изучения в таких условиях взаимодействия не только удаленных нейронов, но и нейронов, расположенных заведомо в пределах рабочего ансамбля. В эксперименте этот вопрос мог бы быть решенным за счет введения ряда микроэлектродов с заданным и фиксированным расположением между ними, однако до последнего времени такой прием в клинических исследованиях является неоправданным, несмотря на работы в области зрительного и слухового протезирования (S imm ons et al., 1965; Brindley, Lewin, 1968; Brindley, 1970; Michelson, 1971). He исключено также, что для нейрофизиологического анализа это не представит особых преимуществ, хотя данный вопрос лучше оставить пока открытым. Проблема чистоты получаемых данных решается нами иначе - применением в исследованиях мозга человека электродов, у которых возможно произвольно варьировать рабочую поверхность и, таким образом, одновременно или последовательно записывать большее или меньшее количество нейронов (Матвеев, Киселев, 1978). Взаимодействие нейронов в пределах одной зоны мозга и между разными его зонами определялось первоначально путем сравнения активности, ее текущей частоты предварительно дискриминированных (способом оконного различения и др.) нейронов, причем основным приемом служил факторный анализ, а эпоха анализа избиралась в соответствии с задачами и плотностью выделенной активности.
Далее с этой целью применялись и другие приемы, и прежде всего изучение регулярности и связности импульсного потока в форме разных видов интервального анализа и прием когерентной детекции - выявления синхронности событий в
удаленных зонах мозга (Бундзен идр., 19756; Кропотов, 1976, 1978; Перепелкин,
1977; Бундзен, Давид, 1979). Эти исследования проводились с целью уточнения не только взаимодействия между нервными элементами, но и всех перестроек местной активности нейронных популяций, характерных для исследуемого вида деятельности. Стимулирующими факторами данного направления работ служили следующие.
1. Экспериментальные исследования, в которых описывалась регулярность и связность импульсных потоков в активности отдельных нейронов и нейронных популяций и возможность развития микроструктурных особенностей паттернов межимпульсных интервалов - узоров межимпульсных разрядов (Gerstein, 1960; Poggio, Vernstein, 1964; Kiang, 1965; Nakahama, 1966; Perkel et al., 1967;. Perkel, Bullok, 1968; Segundo, Perkel, 1969; Verzeano et al., 1970; Василевский и др., 1972; Babb et al., 1973; Василевский, 1976; Nakahama et al., 1977; Толкунов, 1978; Чораян, 1978; Шульгина, 1978, и многие другие).
2. Специальное изучение чисто частотного кодирования, проведенное Эхорном с соавторами (Echorn et al., 1976), показавшее, что при этом виде кодирования возможные потери информации могут быть очень велики - от 10 до 95 %!
3. Данные о функциональных и морфологических основах возможности кодирования появлением специфических узоров импульсной активности (Anderson et al., 1964; Schlag, Villabanca, 1968; Gerstein, Perkel, 1969; Emmers, 1970, 1976; Moore et al., 1970; Sanderson et al., 1973; Katayama, Murata, 1974; Baldissera, Parmiggiani, 1976; Толкунов, 1978).