Лекция 6. Взаимодействие неаллельных генов.
План занятия.
1. Взаимодействие неаллельных генов.
2. Комплементарность.
3. Эпистаз.
4. Полимерия.
5. Плейотропное действие генов
6. Генотип — система генов.
1. Взаимодействие неаллельных генов. Очень редко развитие того или иного признака определяется одним геном. Часто один ген обусловливает развитие нескольких признаков. Такое явление называется плейотропным действием гена. Распространено и обратное — один признак контролируется несколькими генами.
Неаллельные гены — это гены, расположенные в различных участках (локусах) хромосом и кодирующие неодинаковые белки. Неаллельные гены могут взаимодействовать между собой, когда один признак проявляется под действием совокупности нескольких генов.
Первый случай неаллельного взаимодействия был описан в качестве примера отклонения от законов Менделя английскими учеными У. Бетсоном и Р. Пеннетом в 1904 г. при изучении наследования формы гребня у кур.
Различные породы кур характеризуются разной формой гребня:
-Виандотты - низкий, правильный, покрытый сосочками гребень, известный под названием “розовидного”;
-Брамы и некоторые бойцовые куры - узкий и высокий гребень с тремя продольными возвышениями — “гороховидный”.
-Леггорны - простой или листовидный гребень, состоящий из одной вертикальной пластинки. Гибридологический анализ показал, что простой гребень ведет себя как полностью рецессивный признак по отношению к розовидному и гороховидному.
При скрещивании же между собой рас с розовидным и гороховидным гребнем у гибридов первого поколения развивается совершенно новая форма гребня, напоминающая половинку ядра грецкого ореха, в связи с чем гребень был назван “ореховидным”. При анализе второго поколения было установлено, что соотношение разных форм гребня в F2соответствует формуле 9: 3: 3: 1, что указывало на дигибридный характер скрещивания. Была разработана схема скрещивания, объясняющая механизм наследования этого признака.
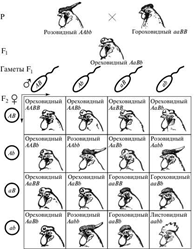 В определении формы гребня у кур принимают участие два неаллельных гена. Доминантный ген R контролирует развитие розовидного гребня, а доминантный ген P — гороховидного. Комбинация рецессивных аллелей этих генов rrpp вызывает развитие простого гребня. Ореховидный гребень развивается при наличии в генотипе обоих доминантных генов.
В определении формы гребня у кур принимают участие два неаллельных гена. Доминантный ген R контролирует развитие розовидного гребня, а доминантный ген P — гороховидного. Комбинация рецессивных аллелей этих генов rrpp вызывает развитие простого гребня. Ореховидный гребень развивается при наличии в генотипе обоих доминантных генов.
Наследование генов, определяющих форму гребня у кур, полностью укладывается в схему дигибридного скрещивания, так как они ведут себя при распределении независимо. Отличие от обычного дигибридного скрещивания проявляется только на уровне фенотипа и сводится к следующему:
1. Гибриды F1не похожи ни на одного из родителей и обладают новым признаком;
2. В F2появляются два новых фенотипических класса, которые являются результатом взаимодействия либо доминантных (ореховидный гребень), либо рецессивных (простой гребень) аллелей двух независимых генов.
Наследование формы гребня у кур можно отнести к к омплементарному взаимодействию неаллельных генов.
2. Комплементарность. К комплементарным, или дополняющим друг друга, генам относятся такие неаллельные гены, которые при совместном проявлении обусловливают развитие нового признака. Действие же каждого из генов в отдельности воспроизводит признак одного из родителей.
Механизм комплементарного взаимодействия подробно изучен на примере наследования окраски глаз у дрозофилы.
Другой пример комплеменетарного действия генов наблюдается при скрещивании двух разновидностей фигурной тыквы со сферической формой плода гибриды первого поколения обладают новым признаком — плоскими или дисковидными плодами. При скрещивании гибридов между собой в F2наблюдается расщепление в соотношении: 9 дисковидных: 6 сферических: 1 удлиненная (9:6:1)
 Анализ схемы показывает, что в определении формы плода принимают участие два неаллельных гена с одинаковым фенотипическим проявлением (сферическая форма). Взаимодействие доминантных аллелей этих генов дает дисковидную форму, взаимодействие рецессивных аллелей — удлиненную.
Анализ схемы показывает, что в определении формы плода принимают участие два неаллельных гена с одинаковым фенотипическим проявлением (сферическая форма). Взаимодействие доминантных аллелей этих генов дает дисковидную форму, взаимодействие рецессивных аллелей — удлиненную.
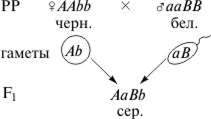
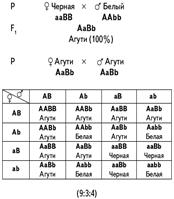
Еще один пример комплементарного взаимодействия дает наследование окраски шерсти у мышей. Дикая серая окраска определяется взаимодействием двух доминантных генов. Ген А отвечает за присутствие пигмента, а ген В — за его неравномерное распределение. Если в генотипе присутствует только ген А (А-bb), то мыши равномерно окрашены в черный цвет. Если присутствует только ген В (ааВ-), то пигмент не вырабатывается и мыши оказываются неокрашенными, так же как и гомозиготный рецессив ааbb. Такое действие генов приводит к тому, что в F2расщепление по фенотипу соответствует формуле 9: 3: 4.
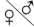
| AB | Ab | aB | ab |
| AB | AABB сер. | AABb сер. | AaBB сер. | AaBb сер. |
| Ab | AABb сер. | AAbb черн. | AaBb сер. | Aabb черн. |
| aB | AaBB сер. | AaBb сер. | aaBB бел. | aaBb бел. |
| ab | AaBb сер. | Aabb черн. | aaBb бел. | aabb бел. |
F2: 9 сер.: 3 черн.: 4 бел.
Комплементарное взаимодействие описано также при наследовании окраски цветов у душистого горошка. Большая часть сортов этого растения имеет пурпурные цветы с фиолетовыми крыльями, которые характерны для дикой сицилийской расы, но есть также сорта с белой окраской. Скрещивая растения с пурпурной окраской цветов с растениями с белыми цветами Бетсон и Пеннет установили, что пурпурная окраска цветов полностью доминирует над белой, и в F2наблюдается соотношение 3: 1. Но в одном случае от скрещивания двух белых растений получилось потомство, состоящее только из растений с окрашенными цветами. При самоопылении растений F1было получено потомство, состоящее из двух фенотипических классов: с окрашенными и неокрашенными цветами в соотношении 9/16: 7/16.
Полученные результаты объясняются комплементарным взаимодействием двух пар неаллельных генов, доминантные аллели которых (С и Р) в отдельности не способны обеспечить развитие пурпурной окраски, так же как и их рецессивные аллели (ссрр). Окраска проявляется только при наличии в генотипе обоих доминантных генов, взаимодействие которых обеспечивает синтез пигмента.
Объяснение такого результата состоит в том, что каждый из доминантных генов не может вызвать появление окраски, определяемой пигментом антоцианом. У душистого горошка есть ген А, обусловливающий синтез бесцветного предшественника пигмента — пропигмента. Ген В определяет синтез фермента, под действием которого из пропигмента образуется пигмент. Цветки душистого горошка с генотипом ааВВ и ААbb имеют белый цвет: в первом случае есть фермент, но нет пропигмента, во втором — есть пропигмент, но нет фермента, переводящего пропигмент в пигмент.
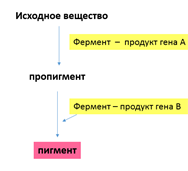 В приведенном примере формула расщепления в F2— 9: 7 обусловлена отсутствием у доминантных аллелей обоих генов собственного фенотипического проявления. Однако такой же результат получается и в том случае, если взаимодействующие доминантные гены имеют одинаковое фенотипическое проявление.
В приведенном примере формула расщепления в F2— 9: 7 обусловлена отсутствием у доминантных аллелей обоих генов собственного фенотипического проявления. Однако такой же результат получается и в том случае, если взаимодействующие доминантные гены имеют одинаковое фенотипическое проявление.
Например, при скрещивании двух сортов кукурузы с фиолетовой окраской зерновок в F1все гибриды имеют желтые зерновки, а в F2наблюдается расщепление 9/16 желт.: 7/16 фиол.
Таким образом, с позиции фенотипического проявления (расщепления по фенотипу в F2) можно выделить 4 варианта комплементарного взаимодействия генов.
1) 9: 3: 3: 1- каждый из доминантных аллелей комплементарных генов в отдельности воспроизводит свой специфический признак (фенотип), а при совместном их присутствии в генотипе проявляется новый признак.
2) 9: 6: 1 — каждый доминантный аллель из комплементарных генов в отдельности воспроизводит один и тот же признак, а вместе они обусловливают проявление нового признака.
3) 9: 3: 4 - один из генов имеет собственное фенотипическое проявление, а действие другого может проявиться только в присутствии первого,что приводит к формированию нового признака.
4) 9: 7 - каждый из комплементарных генов в отдельности не может обусловить развитие признака, но при сочетании в генотипе двух доминантных аллелей этих генов признак проявляется. При этом в F2 расщепление по фенотипу 9:7.
Эпистаз
Эпистаз — взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый — гипостатичным. Если эпистатичный ген не имеет собственного фенотипического проявления, то он называется ингибитором и обозначается буквой I.
Эпистатическое взаимодействие неаллельных генов может быть доминантным и рецессивным.
Доминантный эпистаз
При доминантном эпистазе проявление гипостатичного гена (В, b) подавляется доминантным эпистатичным геном (I > В, b). Расщепление по фенотипу при доминантном эпистазе может происходить в соотношении 12:3:1, 13:3. Последний вариант возникает, когда рецессивная гомозигота по гипостатичному гену фенотипически неотличима от фенотипа I (например, обе формы неокрашены).
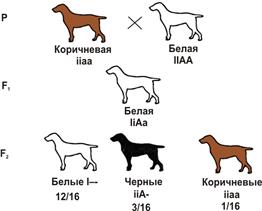
Наследование окраски шерсти у собак (пример доминантного эпистаза):
A — черная окраска, а — коричневая, I — подавляет окраску, i — не подавляет.
Схема наследования окраски шерсти у лошадей
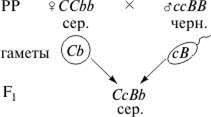
F2
| CB | Cb | cB | cb |
| CB | CCBB сер. | CCBB сер. | CcBB сер. | CcBb сер. |
| Cb | CCBb сер. | CCbb сер. | CcBb сер. | Ccbb сер. |
| cB | CcBB сер. | CcBb сер. | ccBB черн. | ccBb черн. |
| cb | CcBb сер. | Ccbb сер. | ccBb черн. | ccbb рыж. |
F2: 12 сер.: 3 черн.: 1 рыж.
Из схемы видно, что доминантный ген серой окраски С является эпистатичным по отношению к доминантному гену В, который обусловливает черную окраску. В присутствии гена С ген В своего действия не проявляет, и поэтому гибриды F1несут признак, определяемый эпистатичным геном. В F2класс с обоими доминантными генами сливается по фенотипу (серая окраска) с классом, у которого представлен только эпистатичный ген (12/16). Черная окраска проявляется у 3/16 гибридных потомков, в генотипе которых отсутствует эпистатичный ген. В случае гомозиготного рецессива отсутствие гена-супрессора позволяет проявиться рецессивному гену с, который вызывает развитие рыжей окраски.
Доминантный эпистаз описан также при наследовании окраски пера у кур. Белый цвет оперенья у кур породы леггорнов доминирует над окрашенным черных, рябых и других цветных пород. Однако белая окраска других пород (например, плимутроков) рецессивна по отношению к цветному оперению. Скрещивания между особями с доминантной белой окраской и особями с рецессивной белой окраской в F1дают белое потомство. В F2наблюдается расщепление в соотношении 13: 3.
Анализ схемы показывает, что в определении окраски пера у кур принимают участие две пары неаллельных генов. Доминантный ген одной пары (I) является эпистатичным по отношению к доминантному гену другой пары, вызывающему развитие окраски (C). В связи с этим окрашенное оперение имеют только те особи, в генотипе которых присутствует ген С, но отсутствует эпистатичный ген I. У рецессивных гомозигот ссii отсутствует эпистатичный ген, но у них нет гена, который обеспечивает выработку пигмента (C), поэтому они имеют белую окраску.
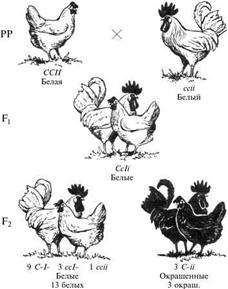
С позиции фенотипического проявления (расщепления по фенотипу в F2) выделено 2 варианта доминантного эпистаза:
1) 12: 3: 1 — форма аавв фенотипически отличается от форм с домининтными аллелями двух генов А_В_ и доминантным аллелем одного гена А_вв, ааВ_.
2)13: 3 – в этом случае рецессивная гомозигота (аавв) фенотипически не отличима от одного из гетерозиготных классов А_вв или ааВ_
Рецессивный эпистаз
Рецессивный эпистаз — это подавление рецессивным аллелем эпистатичного гена в гомозиготном состоянии аллелей гипостатичного гена (ii > В, b). Расщепление по фенотипу может идти в соотношении 9:3:4, 9:7. Последний вариант возникает, когда рецессивная гомозигота по гипостатичному гену фенотипически неотличима от фенотипа ii (например, обе формы неокрашены).
Рецессивный эпистаз проявляется при наследовании окраски шерсти у домовых мышей.
А —окраска агути (рыжевато-серая)
а — черная окраска
В — способствует проявлению окраски
b — супрессор (подавляет действие А и а)
Мыши с генотипом А-bb и ааbb имеют одинаковый фенотип — все белые.
В качестве примера рецессивного эпистаза можно рассмотреть ситуацию с геном альбинизма у животных. Присутствие в генотипе двух аллелей гена альбинизма (аа) не дает возможности проявиться доминантному гену окраски (B) — генотипы ааВ-.
Таким образом выделяют 2 варианта расщипления признаков при рецессивном эпистазе:
1) 9: 3: 4 — 3 варианта проявления фенотипа.
2) 9: 7 — рецессивная гомозигота (аа) по гипостатичному гену фенотипически неотличима от фенотипа ii (например, обе формы неокрашены).
Полимерия
Полимерный тип взаимодействия был впервые установлен Г. Нильсеном-Эле при изучении наследования окраски зерна у пшеницы.
Полимерия — взаимодействие неаллельных генов, при котором степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного гена имеют одинаковый нижний индекс.
Например, у пшеницы А — тёмно-красный цвет зёрен, а — белый цвет зёрен. За цвет отвечают два гена — 1 и 2. Первый ген может быть представлен аллелями А1 и а1, второй — аллелями А2 и а2.
В зависимости от того, каких аллелей больше — А или а, оттенок зёрен будет изменяться.
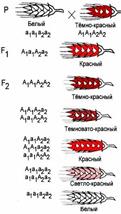
Схема наследования окраски зерна у пшеницы
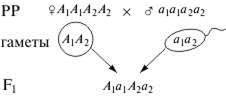
F2
| A1A2 | A1a2 | a1A2 | a1a2 |
| A1A2 | A1A1A2A2 красн. | A1A1A2Aa2 ярко-розов. | A1a1A2A2 ярко-розов. | A1a1A2a2 розов. |
| A1a2 | A1A1A2a2 ярко-розов. | A1A1a2a2 розов. | A1a1A2a2 розов. | A1a1a2a2 бледно-розов. |
| a1A2 | A1a1A2A2 ярко-розов. | A1a1A2a2 розов. | a1a1A2A2 розов. | a1a1A2a2 бледно-розов. |
| a1a2 | A1a1A2a2 розов. | A1a1a2a2 бледно-розов. | a1a1A2a2 бледно-розов. | a1a1a2a2 бел. |
F2: 15 окраш.: 1 бел.
Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным.
При кумулятивной (накопительной) полимерии степень проявления признака зависит от числа доминантных аллелей всех генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак (например, окрас семян пшеницы).
При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов (Резус — фактор). Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление по фенотипу происходит в соотношении 15:1 для двух генов, 63:1 для трёх генов и т.д. Например, цвет кожи у людей зависит от четырёх генов, поэтому от брака двух мулатов практически никогда не рождаются дети с белой кожей.
Плейотропное действие генов.
К взаимодействию неаллельных генов относят также явление плейотропии — множественного действия гена, влияния его на развитие нескольких признаков.
У дрозофилы ген белого цвета глаз одновременно влияет на цвет тела, длины, крыльев, строение полового аппарата, снижает плодовитость, уменьшает продолжительность жизни. У человека известна наследственная болезнь - арахнодактилия ("паучьи пальцы"-очень тонкие и длинные пальцы), или болезнь Марфана. Ген, отвечающий за эту болезнь, вызывает нарушение развития соединительной ткани и одновременно влияет на развитие нескольких признаков: нарушение строения хрусталика глаза, аномалии в сердечно-сосудистой системе.
Схема наследования платиновой окраски у лис
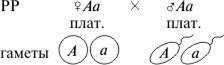
F1
| A | a |
| A | AA погибают | Aa платин. |
| a | Aa платин. | aa черн. |
F1: 2 плат.: 1 черн.