Полное доминирование — вид взаимодействия, при котором в фенотипе гетерозигот присутствует продукт только одного (доминантного) гена и фенотип гетерозигот имеет такое же значение, как фенотип гомозигот по доминантному признаку.
Неполное доминирование — вид взаимодействия, при котором фенотип гетерозигот отличается от фенотипов гомозигот по доминантному и рецессивному признакам и имеет промежуточное значение между ними.
Кодоминирование – В случае кодоминирования у гетерозиготных особей полностью проявляются оба аллельных гена. Классическим примером кодоминирования является взаимодействие генов у человека с кровью четвертой группы (по системеАB0).
Группы крови 0, А, В и АВ определяются геном I. Известны три разновидности этого гена: I A, I B, I 0. Аллельные гены I A и I B полностью доминируют над I 0, а между собой взаимодействуют по типу кодоминирования.
Ген I A обусловливает наличие на мембранах эритроцитов антигена А, ген I B определяет наличие другого антигена — В. Следовательно, у людей с генотипами I A I A и I A I 0 эритроциты несут только антиген А — это кровь второй (А) группы. У людей с генотипами I В I В и I В I 0 красные кровяные клетки содержат только антиген В — это кровь третьей (В) группы. Человек с генотипом I 0 I 0 имеет кровь первой (0) группы — на поверхности его эритроцитов отсутствуют антигены А и В.
У гетерозигот с генотипом I A I B гены I A и I B не подавляют проявление друг друга, а кодоминируют. Каждый из них в полной мере выполняет свою функцию, поэтому эритроциты содержат оба антигена (А и В). В результате этого возникает качественно новый признак — кровь четвертой группы.
Сверхдоминирование – это такое взаимодействие аллельных генов, при котором в гетерозиготном состоянии доминантный аллель проявляет себя намного ярче, чем у доминантной гомозиготы. Этот механизм взаимодействия генов лежит в основе гетерозиса.
Наиболее известный пример Сверхдоминирование — взаимоотношения между нормальным (S) и мутантным (s) аллелями гена, контролирующего структуру гемоглобина у человека. Люди, гомозиготные по мутантной аллели (ss), страдают тяжёлым заболеванием крови — серповидноклеточной анемией
, от которого они гибнут обычно в детском возрасте (эритроциты больного имеют серповидную форму и содержат гемоглобин, структура которого незначительно изменена в результате мутации). Однако в тропической Африке и других районах, где распространена малярия, в популяциях человека постоянно присутствуют все три генотипа — SS, Ss и ss (20—40% населения гетерозиготы — Ss). Оказалось, что сохранение в популяциях человека летальной (смертельной) аллели (s) обусловлено тем, что гетерозиготы (Ss) более устойчивы к малярии, чем гомозиготы по нормальному гену (SS), и, следовательно, обладают отборным преимуществом.
2.Взаимодействие неаллельных генов
Признаки, наследование которых мы рассматривали ранее, контролируются генами одной аллельной пары. Однако многие признаки живых организмов определяются взаимодействием двух или более пар генов, т. е. неаллельными генами. Такое явление получило название взаимодействие неаллельных генов. Основными типами взаимодействия неаллельных генов являются комплементарность, эпистаз и полимерия.
Комплементарными называют гены, обусловливающие при совместном сочетании в генотипе в гомозиготном или гетерозиготном состоянии новое фенотипическое проявление признака. Комплементарность - взаимодействие неаллельных генов, дающее при совместном сочетании в генотипе в гомозиготном или гетерозиготном состоянии новое фенотипическое проявление признака.
Комплементарность выражается в том, что доминантные гены разных аллелей совместно обусловливают появление нового признака, не определявшегося ни одним из этих генов по отдельности. Например, у льна доминантный ген одной аллели (А) определяет розовую окраску цветков, а доминантный ген другой аллели (В) — белую. Если в генотипе растения присутствуют оба доминантных аллеля, его цветки имеют голубую окраску. Таким образом, неаллельные гены А и В действуют комплементарно (взаимно дополняя друг друга), что приводит к формированию нового признака. Суть такого взаимодействия можно отразить в виде схемы.

Как видно из схемы, синтез голубого пигмента из неокрашенного предшественника (пропигмента) осуществляется в два этапа, каждый из которых катализируется особым ферментом. Присутствие этих ферментов в клетках лепестков льна определяется наличием в генотипе соответствующих генов — А и В. Так, ген А определяет наличие фермента, который превращает пропигмент в промежуточный продукт — пигмент розового цвета. Ген В обусловливает присутствие второго фермента, синтезирующего голубой пигмент из розового. Поэтому у растений, имеющих оба доминантных аллеля, в лепестках образуется конечный продукт — пигмент голубого цвета. Если отсутствует ген В (а значит, и соответствующий фермент), в клетках протекает лишь первая стадия — образование розового пигмента. В случае отсутствия гена А (или обоих доминантных аллелей) не осуществляется даже первый этап синтеза пигмента, и лепестки остаются белыми.
Рассмотрим скрещивание дигомозиготных растений льна, имеющих розовые (AAbb) и белые (aaBB) цветки (рис. 33-1.1). В первом поколении наблюдается единообразие гибридов. Все они имеют голубые цветки, что обусловлено наличием в генотипе как гена А, так и гена В. Во втором гибридном поколении происходит расщепление в соотношении 9 голубых: 3 розовых: 4 белых.

Комплементарное взаимодействие генов лежит в основе проявления ряда признаков у различных организмов. Так, по типу комплементарности взаимодействуют гены, контролирующие окраску коконов у тутового шелкопряда и цветков у душистого горошка, форму гребня у кур, цвет оперения у попугаев и т. д. В зависимости от особенностей фенотипического проявления генов в потомстве от скрещивания дигетерозигот могут наблюдаться различные варианты расщепления — 9: 7, 9: 3: 4, 9: 6: 1, 9: 3: 3: 1.
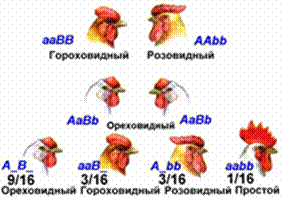
Рис. 6. Наследование формы гребня у кур.
Полимерия. Скрещивая белую и пурпурную фасоли, Мендель столкнулся с явлением полимерии.
Полимерией называют влияние двух, трех и более неаллельных генов на развитие одного и того же признака. Такие гены называют полимерными, или множественными, и обозначают одной буквой с соответствующим индексом, например, А1, А2, а1, а2. Полимерные гены контролируют большинство количественных признаков организмов: высоту растения, массу семян, масличность семян, содержание сахара в корнеплодах сахарной свеклы, удойность коров, яйценоскость, вес тела и т.д.
Известны две разновидности полимерии.
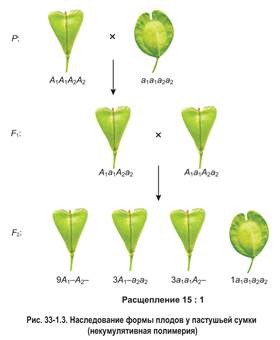
При некумулятивной полимерии доминантный признак проявляется в полной мере при наличии в генотипе хотя бы одного доминантного гена. Иными словами, количество доминантных генов не влияет на степень выраженности признака. Так, у пастушьей сумки форма плодов определяется двумя парами полимерных генов. При этом рецессивный признак — овальные плоды проявляется только у растений, имеющих генотип а1а1а2а2. Если в генотипе присутствует хотя бы один доминантный ген (неважно какой именно — А1 или А2), растение обладает доминантным признаком — плодами треугольной формы.
Скрещивание доминантной и рецессивной дигомозигот (рис. 33-1.3) приводит к появлению в первом поколении дигетерозиготных гибридов с треугольными плодами (А1а1А2а2). Во втором поколении наблюдается расщепление в соотношении 15: 1.
В случае кумулятивной полимерии степень выраженности доминантного признака определяется количеством доминантных генов в генотипе. Чем больше таких генов, тем сильнее проявляется данный признак. Например, у пшеницы определенных сортов окраска зерен контролируется тремя парами неаллельных генов. Растения с генотипом а1а1а2а2а3а3 имеют белые зерна, а обладающие генотипом А1А1А2А2А3А3 — темно-красные. У растений с другими генотипами наблюдаются различные промежуточные варианты окраски, зависящие от числа доминантных генов. Так, наличие в генотипе только одного доминантного гена обусловливает бледно-розовый цвет зерен, двух — розовый, трех — светло-красный и т. д.
Установлено, что по типу кумулятивной полимерии наследуются многие количественные признаки организмов: рост человека, цвет его кожи (т. е. количество меланина), яйценоскость кур, молочность коров, содержание сахарозы в корнеплодах сахарной свеклы, белков в эндосперме семян злаков и др.
У человека по типу полимерии наследуется цвет кожи. У негров темная окраска кожи определяется 2 парами неаллельных генов А1А1А2А2, у белого человека – соответственно а1а1а2а2. От брака негра и белой женщины родится средний мулат с генотипом А1а1А2а2. От брака средних мулатов вероятность рождения нега – 1/16, темного мулата (три доминантные аллели) – 4/16, среднего мулата (две доминантные аллели) – 6/16; светлого мулата (одна доминантная аллель) – 4/16, белого – 1/16.
Эпистаз — тип взаимодействия, при котором ген одной аллели подавляет проявление генов другой аллельной пары. Такие гены, блокирующие фенотипическое проявление других неаллельных генов, называются эпистатическими, генами-ингибиторами или супрессорами. Подавляемые гены, в свою очередь, получили название гипостатических. В зависимости от того, доминантным или рецессивным является ген-ингибитор, различают доминантный и рецессивный эпистаз.
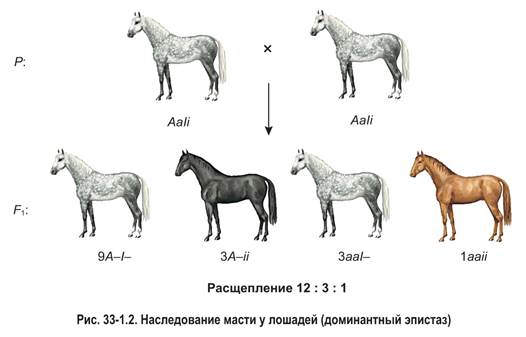
Примером доминантного эпистаза может служить взаимодействие двух аллелей, определяющих масть (окраску) лошадей. Так, доминантный ген одной аллельной пары (А) обусловливает вороную (черную) масть лошади, а рецессивный ген (а) — рыжую. Однако эти гены могут проявиться фенотипически только при отсутствии в генотипе лошади доминантного гена другой аллели — ингибитора (I). При наличии ингибитора проявление генов вороной и рыжей масти подавляется. В рассматриваемом примере ген-супрессор имеет собственное фенотипическое проявление: в случае его присутствия у особей развивается серая окраска.
Рассмотрим скрещивание дигетерозиготных лошадей (рис. 33-1.2). Они имеют серую масть, что обусловлено наличием в генотипе доминантного гена-супрессора (I). В потомстве от скрещивания наблюдается расщепление 12: 3: 1. Все особи, содержащие в генотипе эпистатический ген (9A—I— и 3aaI—), наследуют серую масть. У жеребят, не имеющих гена I, проявляется вороная либо рыжая окраска в соотношении 3 вороные (3A–ii) к 1 рыжей (1aaii).
Если ген-супрессор является рецессивным, то взаимодействие неаллельных генов протекает по типу рецессивного эпистаза (криптомерии). Такое взаимодействие наблюдается, например, при наследовании цвета семян фасоли. У этого растения красная окраска семян (А) доминирует над желто-коричневой (а). Проявление рецессивного ингибитора (i) ведет к блокировке действия генов красной и желто-коричневой окраски, и семена оказываются белыми. Однако действие рецессивного эпистатического гена может проявиться только при отсутствии в генотипе соответствующего ему доминантного аллеля (I). В случае наличия такого гена действие супрессора подавляется. Так, скрещивание дигетерозиготных растений фасоли приводит к расщеплению среди потомков в соотношении: 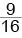 с красными,
с красными, 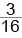 с желто-коричневыми и
с желто-коричневыми и 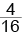 с белыми семенами.
с белыми семенами.

Эпистаз наблюдается при наследовании многих признаков живых организмов. При различных формах эпистаза и разных особенностях проявления генов расщепление в потомстве от скрещивания дигетерозиготных особей может составлять 13: 3, 12: 3: 1 или 9: 3: 4.
Множественный аллелизм.
Окраска семян гороха, наследование которой изучал Г. Мендель, определяется двумя аллелями — доминантным и рецессивным. Однако многие гены представлены не двумя, а бóльшим числом аллелей. Например, ген, определяющий у человека группы крови (по системе АВ0), как уже отмечалось, существует в трех формах — I A, I B, I 0.
Явление существования гена более чем в двух аллельных формах называется множественным аллелизмом. При этом у любого организма, соматические клетки которого содержат диплоидный набор хромосом, имеется не более двух аллелей каждого гена.
Множественные аллели нередко образуют серии, в которой каждый предыдущий аллель доминантен по отношению к последующим. Так, ген, определяющий цвет шерсти кролика, представлен несколькими аллельными формами (рис.14.9 в учебнике на стр. 178).
При этом аллель С, обусловливающий черную окраску, доминирует над аллелями шиншилловой (с ch), гималайской (с h) и белой (с а) окраски. Ген, контролирующий шиншилловую окраску, доминирует над генами, определяющими гималайскую и белую. И наконец, аллель с h доминантен по отношению к аллелю с а. Это можно выразить в виде: С > с ch > с h > с а. Подобным образом взаимодействуют гены, контролирующие окраску глаз у плодовой мушки дрозофилы (красная > коралловая > вишневая > абрикосовая и т. д. до белой), форму белых пятен на листьях клевера ползучего и др.
Но один ген может влиять и на несколько признаков, это явление получило название плейотропия, или множественное действие гена. Множественное действие гена имеет биохимическую природу: один белок-фермент, образующийся под контролем одного гена, определяет развитие не только данного признака,нои воздействует на развитие других признаков и свойств, вызывая их изменение.
Впервые было обнаружено Г.Менделем, который обнаружил, что у растений с пурпурными цветками всегда имелись красные пятна в пазухах листьев, а семенная кожура была серого или бурого цвета. То есть развитие этих признаков определяется действием одного наследственного фактора (гена).
У дрозофилы окраска глаз определяется доминантным геном W, рецессивная аллель w в гомозиготном или гемизиготном состоянии приводит к развитию белоглазости. Но кроме этого, белоглазые особи имеют более светлую окраску внутренних органов, меньшую плодовитость и меньшую продолжительность жизни. У человека ген, определяющий рыжий цвет волос, обуславливает более светлую кожу и появление веснушек.
Таким образом, один ген может определять развитие одного признака; один ген может влиять на проявление нескольких признаков (плейотропия); за развитие одного признака могут отвечать несколько генов (полимерия), то есть генотип – это не просто сумма генов, а система взаимодействующих генов. Кроме того, на проявление действия генов влияют и условия окружающей внешней среды. Следовательно, генотип является системой генов, взаимодействующих между собой и факторами среды обитания.
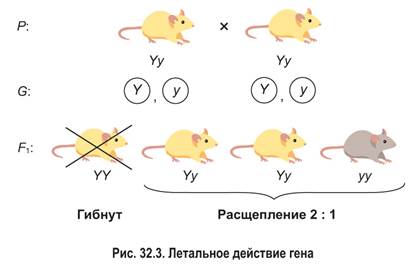
Летальное действие генов. У ряда организмов обнаружены гены, которые в гомозиготном состоянии вызывают нарушения индивидуального развития, приводящие к гибели особей на ранних этапах онтогенеза (чаще всего в эмбриональном периоде). Такие гены называются летальными.
Например, у мышей летальным действием обладает доминантный ген Y, определяющий желтый цвет шерсти. Рецессивный ген y обусловливает серую окраску. Гомозиготные эмбрионы с генотипом YY погибают еще до рождения, а гетерозиготные мышата Yy являются жизнеспособными и имеют желтую шерсть. Поэтому при скрещивании желтых мышей (они могут быть только гетерозиготами) в потомстве наблюдается расщепление 2 желтые: 1 серая, а не 3: 1 (рис. 32.3).
У карпа летальное действие имеет ген, определяющий недоразвитие чешуи. Гомозиготные особи гибнут до выхода из икринок, а гетерозиготные выживают. Такие карпы имеют чешую лишь на боковой линии тела, поэтому их называют линейными. Действие летального гена, обусловливающего укорочение клюва у кур, приводит к тому, что гомозиготные цыплята оказываются неспособными вылупиться из яиц. Клюв этих цыплят так мал, что они не могут пробить скорлупу и поэтому погибают. У человека известен доминантный ген, вызывающий у гомозиготных эмбрионов несовместимые с жизнью нарушения развития скелета. Гетерозиготные организмы выживают, у этих людей наблюдается укорочение средних фаланг пальцев — брахидактилия.*
Некоторые гены обладают плейотропным (множественным) действием, т. е. контролируют не один признак, а несколько. Явление плейотропии объясняется тем, что один и тот же белок — продукт такого гена — может входить в состав разных тканей и (или) участвовать в различных биохимических процессах.
Например, у человека рецессивный ген, обусловливающий рыжий цвет волос, также определяет появление веснушек и окраску кожи, более светлую, чем у других людей. Доминантный ген, вызывающий развитие аномально длинных пальцев кистей и стоп («паучьих пальцев»), обусловливает дефект хрусталика глаза, пороки сердца и некоторые другие признаки. Один из генов дрозофилы, контролирующий белую окраску глаз, приводит также к снижению плодовитости особей. У овса один и тот же ген определяет длину остей и окраску чешуй зерновок.