СЛАЙД
Дыхание человека меняется в зависимости от состояния его организма. Оно спокойное, редкое во время сна, частое и глубокое при физических нагрузках, прерывистое. Неровное по время эмоций. При погружении в холодную воду у человека на время останавливается дыхание, «дух захватывает».
Дыхание — совокупность процессов, обеспечивающих потребление организмом кислорода и выделение двуокиси углерода. Поступление кислорода из атмосферы к клеткам необходимо для биологического окисления органических веществ, в результате которого освобождается энергия, необходимая для жизни организма
Физиологическая роль легочного дыхания
Физиологическая роль легочного дыхания состоит в обеспечении оптимального газового состава артериальной крови.
Для нормальной интенсивности процессов тканевого дыхания необходимо, чтобы кровь, поступающая в тканевые капилляры, всегда была насыщена кислородом и не содержала СО, в количествах, препятствующих отдаче его из тканей. Поскольку при прохождении крови через капилляры легких между плазмой и альвеолярным воздухом устанавливается практически почти полное газовое равновесие, то оптимальное содержание газов в артериальной крови определяет соответствующий состав альвеолярного воздуха. Оптимальное содержание газов в альвеолярном воздухе достигается путем изменения объема легочной вентиляции в зависимости от условий, существующих в данный момент в организме.
3 СЛАЙД
Регуляция внешнего дыхания
Регуляция внешнего дыхания представляет собой физиологический процесс управления легочной вентиляцией, который направлен на достижение конечного приспособительного результата — обеспечение оптимального газового состава внутренней среды организма (крови, интерстициальной жидкости, ликвора) в постоянно меняющихся условиях его жизнедеятельности. Управление дыханием осуществляется по принципу обратной связи: при отклонении от оптимальных величин регулируемых параметров (рН, напряжение О, и СО,) изменение вентиляции направлено на их нормализацию. Избыток, например, водородных ионов во внутренней среде организма (ацидоз) приводит к усилению вентиляции, а их недостаток (алкалоз) — к уменьшению интенсивности дыхания. В обоих случаях изменение вентиляции является средством достижения главной цели регуляции дыхания — оптимизации газового состава внутренней среды (прежде всего, артериальной крови).
Регуляция внешнего дыхания осуществляется путем рефлекторных реакций, возникающих в результате возбуждения специфических рецепторов, заложенных в легочной ткани и сосудистых рефлексогенных зонах. Центральный аппарат регуляции дыхания представляют нервные образования спинного мозга, продолговатого мозга и вышележащих отделов нервной системы. Основная функция управления дыханием осуществляется дыхательными нейронами ствола головного мозга, которые передают ритмические сигналы в спинной мозг к мотонейронам дыхательных мышц.
4 СЛАЙД
Дыхательный центр
Дыхательным центром называют совокупность взаимно связанных нейронов центральной нервной системы, обеспечивающих координированную ритмическую деятельность дыхательных мышц и постоянное приспособление внешнего дыхания к изменяющимся условиям внутри организма и в окружающей среде.
Еще в начале XIX века было показано, что в продолговатом мозге на дне IV желудочка в каудальной его части (в области так называемого писчего пера) расположены структуры, разрушение которых уколом иглы ведет к прекращению дыхания и гибели организма. Этот небольшой участок мозга в нижнем углу ромбовидной ямки, жизненно необходимый для поддержания ритмического дыхания, был назван «дыхательным центром». В дальнейшем было показано, что дыхательный центр расположен в медиальной части ретикулярной формации продолговатого мозга, в области obex, вблизи stria acusticae, и состоит из двух отделов:
1. инспираторный отдел («центра вдоха»),
2. экспираторный отдел («центра выдоха»).
5 СЛАЙД
Дыхательные нейроны
В ретикулярной формации продолговатого мозга обнаружены так называемые дыхательные нейроны, одни из которых разряжаются серией импульсов в фазу вдоха, другие — в фазу выдоха. В зависимости от того, каким образом активность дыхательных нейронов коррелирует с фазами дыхательного цикла, их называют инспираторными или экспираторными. В продолговатом мозге не найдено строго обособленных областей, которые содержали бы только инспиратерные или только экспираторные дыхательные нейроны. Тем не менее, инспираторные и экспираторные нейроны рассматривают как две функционально различные популяции, внутри которых нейроны связаны между собой сетью аксонов и синапсов. Исследования активности одиночных нейронов ретикулярной формации продолговатого мозга привели к заключению, что область расположения дыхательного центра не может быть очерчена строго и однозначно. Так называемые дыхательные нейроны обнаружены почти на всем протяжении продолговатого мозга. Однако, в каждой половине продолговатого мозга есть участки ретикулярной формации, где дыхательные нейроны сгруппированы с более высокой плотностью.
Дорсальная группа дыхательных нейронов
Дорсальная группа дыхательных нейронов продолговатого мозга находится вентролатеральнее ядра одиночного пучка и состоит, главным образом, из инспираторных нейронов. Часть этих клеток дает нисходящие пути, идущие, в основном, в составе солитарного тракта и образующие у человека моносинаптические контакты с мотонейронами диафрагмального нерва в передних рогах 3-6 шейных сегментов спинного мозга. Нейроны диафрагмального ядра спинного мозга разряжаются или непрерывно (с учащением, в фазу вдоха) или залпами, подобно активности дыхательных нейронов продолговатого мозга. Движения диафрагмы, обеспечивающие от 70 до 90% дыхательного объема, связаны именно с нисходящими влияниями дорсальной группы инспираторных нейронов продолговатого мозга.
Вентральная группа дыхательных нейронов
Вентральная группа дыхательных нейронов расположена в области обоюдного и ретроамбигуального ядер. Нейроны этой группы посылают нисходящие волокна к мотонейронам межреберных и брюшных мышц. Инспираторные мотонейроны спинного мозга концентрируются, главным образом, во 2- 6, а экспираторные — в 8- 10 грудных сегментах. В вентральной группе нейронов продолговатого мозга находятся также эфферентные преганглионарные нейроны блуждающего нерва, обеспечивающие синхронные с фазами дыхания изменения просвета дыхательных путей. Максимум активности нейронов блуждающего нерва, вызывающей повышение тонуса гладких мышц воздухоносных путей, наблюдается в конце выдоха, а минимум — в конце вдоха.
6 СЛАЙД
Характер ритмической активности дыхательных нейронов
В продолговатом мозге обнаружены дыхательные нейроны с различным характером ритмической активности. Только у части инспираторных и экспираторных нейронов начало разряда и длительность серии импульсов строго совпадают с периодом соответствующей фазы дыхательного цикла, Однако, при всем разнообразии видов возбуждения разных дыхательных нейронов продолговатого мозга у каждого из них характер ритмической активности остается, как правило, постоянным.
На этом основании различают:
а) «Полные » инспираторные и экспираторные нейроны, ритмическое возбуждение которых по времени точно совпадает с соответствующей фазой дыхания;
б) «Ранние » инспираторные и экспираторные нейроны, дающие короткую серию импульсов до начала вдоха или выдоха;
в) «Поздние «, проявляющие залповую активность уже после начала инспирации или экспирации:
г) «Инспираторно- экспираторные «, начинающие возбуждаться в фазе вдоха и остающиеся активными в начале выдоха;
д) «Экспираторно-инспираторные «, активность которых начинается во время вдоха и захватывает начало выдоха;
е) «Непрерывные «, работающие без пауз, но с увеличением частоты импульсов во время вдоха или выдоха (рис.8.9).
 Рис.8.9. Активность различных групп дыхательный нейронов
Рис.8.9. Активность различных групп дыхательный нейронов
Рис.8.9. Активность различных групп дыхательный нейронов продолговатого мозга в связи с фазами дыхательного цикла.I — вдох,II — выдох.
Нейроны:
1 — полные;
2 — ранние;
3 — поздние инспираторные;
4,5,6 — аналогичные экспираторные;
7 — инспираторно-экспираторные;
8 —экспираторно-инспираторные;
9,10 — нейроны, обладающие непрерывной активностью с усилением в различные фазы цикла.
Нейроны каждой разновидности не разбросаны по отдельности и нередко находятся друг от друга на расстояние не более 100 мкм. Полагают, что различные виды дыхательных нейронов образуют своеобразные микрокомплексы, которые служат теми очагами, где формируется автоматизм дыхательного центра. Типичным ритмообразующим комплексом является система из четырех нейронов («ранних» и «поздних» инспираторных и экспираторных), объединенных возвратными связями и способных в совокупности генерировать залповую активность. Каждый цикл начинается с активности «раннего» инспираторного нейрона. Затем возбуждение переходит последовательно на «поздний» инспираторный нейрон, «ранний» и «поздний» экспираторные нейроны и снова на «ранний» инспираторный. Благодаря наличию возвратных связей, нейрон каждой ритмообразующей группы, возбуждаясь, оказывает тормозное воздействие на два предшествующих ему в цикле нейрона. Так называемые «полные» инспираторные и экспираторные нейроны обеспечивают передачу возбуждения по нисходящим путям спинного мозга к мотонейронам, иннервирующим дыхательные мышцы.
После перерезки у экспериментальных животных ствола мозга ниже варолиева моста дыхательные движения сохраняются. Однако, изолированный от нисходящих влияний дыхательный центр способен обеспечить лишь примитивное дыхание, при котором длительный выдох периодически прерывается короткими вдохами. Для стабильности и координации дыхательного ритма, обуславливающей дыхание с плавным характером перехода от вдоха к выдоху, необходимо, в первую очередь, участие нервных образований варолиева моста.
7 СЛАЙД
Пневмотаксический центр
В передней части варолиева моста обнаружена область, названная пневмотаксическим центром, разрушение которой приводит к удлинению фаз вдоха и выдоха, а электрическая стимуляция различных ее зон — к досрочному переключению фаз дыхания. При перерезке ствола мозга на границе между верхней и средней третью варолиева моста и одновременном пересечении обоих блуждающих нервов дыхание останавливается на фазе вдоха, лишь иногда прерываемой экспираторными движениями (так называемый апнейзис). На основании этого был сделан вывод, что дыхательный ритм возникает в результате периодического торможения тонической активности нейронов продолговатого мозга афферентной импульсацией, приходящей по блуждающему нерву и действующей через экспираторные нейроны, а после перерезки блуждающего нерва — вследствие ритмического торможения, поступающего из пневмотаксического центра варолиева моста.
В ростральных отделах варолиева моста, в медиальном парабрахиальном ядре, в участках мозговой ткани вентральнее его, а также в структурах, относящихся к управлению дополнительными дыхательными мышцами, т.е. в том месте, которое идентифицируют как пневмотаксический центр, найдено наибольшее количество дыхательных нейронов моста. В отличие от нейронов продолговатого мозга, стабильно сохраняющих характер залповой активности, в варолиевом мосту один и тот же дыхательный нейрон может изменить характер своей деятельности. Дыхательные нейроны варолиева моста организованы в группы, состоящие из 10-12 нейронов разного вида. Среди них много так называемых переходных (фазово-охватывающих) нейронов, проявляющих с максимумом частоты при смене фаз дыхательного цикла. Этим нейронам приписывают функцию связывания различных фаз дыхательного цикла, подготовки условий для прекращения фазы вдоха и перехода к выдоху. Пневмотаксический центр варолиева моста связан с дыхательным центром продолговатого мозга восходящими и нисходящими проводящими путями. К медиальному парабронхиальному ядру и ядру Келликера-Фузе из продолговатого мозга поступают аксоны нейронов одиночного пучка и ретроамбигуального ядра. Эти аксоны являются основным входом в пневмотаксического центра. Отличительной чертой активности дыхательных нейронов варолиева моста является то, что при нарушении связи с продолговатым мозгом они теряют залповый характер импульсации и модуляцию частоты импульсов в ритме дыхания.
Считается, что пневмотаксический центр получает импульсы от инспираторной части дыхательного центра продолговатого мозга и посылает импульсы обратно к дыхательному центру в продолговатый мозг, где они возбуждают экспираторные и тормозят инспираторные нейроны. Дыхательные нейроны варолиева моста первыми получают сведения о необходимости приспособления дыхания к изменяющимся условиям и соответствующим образом меняют активность нейронов дыхательного центра, а переходные нейроны обеспечивают плавную смену вдоха на выдох. Таким образом, благодаря совместной работе с пневмотаксическим комплексом, дыхательный центр продолговатого мозга может осуществлять ритмическую смену фаз дыхательного цикла с оптимальным соотношением длительности вдоха, выдоха и дыхательной паузы. Однако, для нормальной жизнедеятельности и поддержания адекватного потребностям организма дыхания необходимо участие не только варолиева моста, но и вышележащих отделов головного мозга.
СЛАЙД
Дыхательные движения осуществляются благодаря координированной, согласованной деятельности дыхательных мышц.
Нейроны, дающие начало двигательным нервам дыхательных мышц, расположены в передних рогах спинного мозга. Нейроны грудобрюшного нерва, вызывающие сокращение диафрагмы, расположены у человека в 3-м и 4-м (реже в 5-м) шейных сегментах спинного мозга. Межреберные нервы исходят из грудных сегментов спинного мозга.
Но если отделить спинной мозг от продолговатого мозга, то дыхательные движения прекращаются. Однако они сохраняются, если разрез сделать выше продолговатого мозга. Это доказывает, что согласованная деятельность нейронов двигательных нервов, производящих координированные сокращения дыхательной мускулатуры, вызывается участком нейронов — дыхательным центром, который расположен в продолговатом мозге (Легаллуа, Флуранс
 Возбуждение центра вдоха тормозит центр выдоха, и наоборот. Более возбудимы инспираторные нейроны. На поверхности продолговатого мозга у основания четвертого желудочка расположен центр, регулирующий частоту дыхания.
Возбуждение центра вдоха тормозит центр выдоха, и наоборот. Более возбудимы инспираторные нейроны. На поверхности продолговатого мозга у основания четвертого желудочка расположен центр, регулирующий частоту дыхания.
В варолиевом мосте находятся два центра, регулирующие дыхание: в верхней части моста расположен «центр пневмотаксиса», необходимый для нормального дыхания, а в средней и нижней частях — «апнейстический центр», вызывающий судорожные редкие вдохи.
Если перерезать спинной мозг в шейной части и долго производить искусственное дыхание, то после его прекращения начинаются ритмические сокращения дыхательных мышц. Эти сокращения вызываются ядрами двигательных нервов дыхательных мышц, расположенными в передних рогах спинного мозга. Однако это не нормальные дыхательные движения, так как они не координированы.
Координированные дыхательные движения вызываются только дыхательным центром продолговатого мозга, который подчиняет себе центры спинного мозга.
Дыхательный «автоматизм». Дыхание регулируется: 1) нервно- гуморально благодаря возбуждению нейронов дыхательного центра продолговатого мозга колебаниями химического состава притекающей к ним крови; 2) рефлекторно благодаря притоку афферентных импульсов в дыхательный центр продолговатого мозга.
Дыхательный центр вызывает сокращения дыхательной мускулатуры и после перерезки блуждающих нервов, т. е. после выключения рефлекторной саморегуляции дыхания.
Это происходит благодаря тому, что дыхательный центр одновременно возбуждается не только рефлекторно, но и изменением химического состава притекающей к нему крови. Можно отрезать нее афферентные нервы, а деятельность дыхательного центра не прекратится. Это объясняется тем, что важнейший возбудитель дыхательного центра — внутренний, автоматический, главным образом углекислота, накапливающаяся в крови, а также повышение концентрации водородных ионов при накоплении других кислот.
Дыхательный центр у детей легко возбудим. Дыхание у них значительно учащается при психических возбуждениях, небольших физических упражнениях, незначительном повышении температуры тела и окружающей среды.
В легких имеются механически раздражаемые рецепторы (механорецепторы), которые делятся на: медленно адаптирующиеся, т. е. приспособляющиеся к раздражению, рецепторы растяжения, расположенные в глубине легких, в стенках крупных бронхов и в трахее; быстро адаптирующиеся рецепторы растяжения и спадения легких, находящиеся в глубине легких и в слизистой оболочке бронхов, и рецепторы промежуточной ткани легочных альвеол.
9 СЛАЙД
 Медленно адаптирующиеся рецепторы составляют примерно 2/3, они возбуждаются при вдохе. Частота импульсов возбуждения равна 40 — 100 в 1 с и зависит от объема вдыхаемого воздуха и в малой степени от его состава. При увеличении объема легких частота импульсов возрастает. Афферентные импульсы передаются, по толстым волокнам блуждающих нервов.
Медленно адаптирующиеся рецепторы составляют примерно 2/3, они возбуждаются при вдохе. Частота импульсов возбуждения равна 40 — 100 в 1 с и зависит от объема вдыхаемого воздуха и в малой степени от его состава. При увеличении объема легких частота импульсов возрастает. Афферентные импульсы передаются, по толстым волокнам блуждающих нервов.
Быстро адаптирующиеся рецепторы составляют около Уз, на раздувание легких они реагируют короткой, быстро исчезающей группой импульсов, передаваемых по тонким волокнам блуждающих нервов. Рецепторов спадения легких очень мало и они возбуждаются при нормальном выдохе только при сильном их спадении. I» метро адаптирующиеся рецепторы растяжения, спадения и слизистой бронхов идентичны. Они названы ирритантными.
10 СЛАЙД
Рецепторы трахеи и крупных бронхов отвечают на слабые механические раздражения. Благодаря поступлению афферентных импульсов из легких, плевры и из рецепторов мышц в дыхательный центр из последнего направляются эфферентные импульсы и спинной мозг к ядрам двигательных нервов дыхательных мышц, Которые вызывают их сокращение.
Во время вдоха, когда легкие в достаточной степени растягиваются, возникает механическое раздражение рецепторов в легких и плевре, которое рефлекторно прекращает сокращение дыхательных мышц, участвующих в акте вдоха.
Рефлекторное торможение вдыхательных мышц при увеличении объема легких, уменьшение частоты и силы их сокращений вызывается усилением раздражения медленно адаптирующихся рецепторов. Интенсивность торможения пропорциональна увеличению растяжения легких до полной остановки вдоха. При этом рефлекторно снижается также тонус гладких мышц трахеи и бронхов. При усилении раздувания легких появляется кратковременное, в пределах секунды, возбуждение вдыхательных мышц.
Наоборот, при выдохе, когда легкие достигли известной степени расслабления, раздражение рецепторов в легких и плевре вызывает рефлекторно сокращение вдыхательных мышц. Это сокращение вдыхательных мышц при спадении легких вызывается раздражением быстро адаптирующихся рецепторов. Рецепторы диафрагмы в обычных условиях почти не участвуют в регуляции дыхания. При расширении грудной клетки во время вдоха раздражаются рецепторы кожи, покрывающей грудную клетку, межреберных мышц и мышц брюшной стенки, в которых особенно много рецепторов, что рефлекторно усиливает сокращение вдыхательных мышц. Однако сильное раз-дражение этих рецепторов при значительном расширении грудной клетки тормозит сокращения вдыхательных мышц.
Следовательно, вдох рефлекторно регулирует выдох, а выдох регулирует вдох (теория Геринга — Брейера).
Рефлекторная саморегуляция дыхания имеет защитное значение, так как она препятствует чрезмерному растяжению легких при вдохе. Афферентные импульсы из легких и плевры, поступаю-щие по блуждающим нервам во время вдоха, вызывают торможение вдыхательного центра Такое же торможение вдыхательного центра вызывают афферентные импульсы из вдыхательных мышц.
При перерезке обоих блуждающих нервов дыхание сохраняется, но оно становится более глубоким и редким. Углубление дыхания после перерезки блуждающих нервов происходит благодаря тому, что прерываются афферентные волокна, вызывающие торможение дыхательного центра
Раздражение центрального конца перерезанного блуждающего нерва во время вдоха вызывает угнетение вдыхательных движений задержку дыхания на выдохе.
Еще более резкое торможение вдоха вызывает раздражение центрального конца верхнего гортанного нерва.
Возбудимость дыхательного центра по отношению к рефлекторным и нервно-гуморальным влияниям поддерживается также афферентными импульсами из рецепторов, расположенных вне дыхательного аппарата.
Дыхание рефлекторно изменяется при раздражениях рецепторов кожи (прикосновение, тепло, холод), органов зрения, слуха, обоняния и вкуса. Дыхание изменяется при притоке афферентных импульсов из рецепторов скелетных мышц и сухожилий туловища, рук и ног.
Особо важное защитное значение имеют раздражения слизистых оболочек дыхательных путей. Раздражение пылью в дыхательных путях вызывает судорожные выдыхательные движения при закрытой голосовой щели (кашель).
Когда раздражающие вещества, например пары аммиака, действуют на окончания тройничного нерва в носоглотке, происходит рефлекторная задержка дыхания, при этом может наступить сужение бронхов, которое тоже имеет защитное значение.
Раздражение носоглотки пылью или слизью вызывает чихание — глубокий вдох, а затем очень сильный и быстрый выдох при закрытом рте.
На дыхательный центр влияют также раздражения нервных окончаний в дуге аорты и каротидном синусе. Увеличение кровяного давления в них рефлекторно задерживает дыхание, а уменьшение кровяного давления, наоборот, усиливает дыхание. Рефлекторное возбуждение дыхательного центра вызывается также раздражением хеморецепторов дуги аорты и каротидного синуса углекислотой при повышенном содержании ее в крови и раздражениями рецепторов внутренних органов.
Влияние больших полушарий головного мозга на деятельность дыхательного центра. Наиболее тонкая форма регуляции дыхания — обеспечение соответствия дыхательных движений условиям жизни — осуществляется при участии больших полушарий головного мозга.
Предполагается, что нейроны больших полушарий более возбудимы к повышению напряжения углекислоты в крови, чем дыхательный центр в продолговатом мозге.
Роль недостатка кислорода и избытка углекислоты в крови. Организм никаких запасов кислорода не имеет, кроме того, который поглощен при вдохе.
11 СЛАЙД
Недостаток кислорода в крови (гипоксемия), возникающий при понижении его парциального давления, непосредственно не возбуждает дыхательный центр. Человек не может заметить момент наступления опасной для жизни гипоксемии и теряет сознание, не имея никаких ощущений кислородного голодания (например, при постепенном подъеме на большие высоты, при угаре).
Недостаток кислорода в организме или в отдельных тканях называется гипоксией.
Различают: 1) дыхательную гипоксию при недостатке кислорода во вдыхаемом воздухе или при нарушении регуляции дыхания, 2) циркуляторную — при нарушении кровообращения, 3) анемическую — при недостатке количества крови или при нарушениях ее дыхательной функции и 4) токсическую — при отравлениях некоторыми ядами.
Защитные приспособления организма различны в зависимости от типа гипоксии. Быстрее реагируют на гипоксию большие полушария головного мозга и высшие органы чувств.
Недостаточное снабжение кислородом нейронов дыхательного центра (гипоксия) само по себе не возбуждает дыхательный центр. Гипоксия может вызвать возбуждение дыхательного центра лишь вторично вследствие накопления в нем большого количества Кислот, которые не удаляются благодаря недостаточному снабжению его кислородом. Это накопление кислот сопровождается по- иышением концентрации водородных ионов. Возбуждение дыха- if'ii.noro центра вызывает усиление вентиляции легких, умлич-енное удаление углекислого газа и, следовательно, понижение концентрации водородных ионов в крови.
12 СЛАЙД
Таким образом, дыхательный центр поддерживает постоянство pH в крови. При питании мясной пищей, которая богата кислотами, дыхательный центр возбуждается сильнее, дыхание усиливается, что ведет к ускоренному удалению углекислого газа и сохранению постоянства pH крови.
При питании растительной пищей, которая богата щелочами, возбуждение дыхательного центра снижается, дыхание уменьшается, что замедляет удаление углекислого газа и сохраняет постоянство pH в крови.
Возбуждающее действие углекислого газа на дыхательный центр было доказано в опыте на животных с перекрестным кровообращением.
Опыт Фредерика заключается в том, что под наркозом сонная артерия одной собаки соединяется с сонной артерией другой собаки таким образом, чтобы головной мозг каждой собаки получал кровь из тела другого животного. Если давать вдыхать одной из собак СО2, то вентиляция легких увеличивается не у нее, а у другой собаки. Точно так же удушье одной из собак, сопровождающееся накоплением у нее углекислоты в крови, вызывает усиление дыхания у другой собаки. Этот опыт не оставляет сомнения в том, что дыхательный центр возбуждается увеличением содержания углекислоты в омывающей его крови.
Доказано, что дыхание у людей в герметически замкнутом пространстве вызывает одышку (диспноэ), т. е. учащение и углубление дыхания.
При небольшом увеличении количества С02 во вдыхаемом воздухе содержание его в альвеолярном воздухе благодаря возбуждению дыхательного центра и увеличению вентиляции легких мало изменяется. При большом увеличении количества СО2 во вдыхаемом воздухе даже усиленная вентиляция легких не может возместить увеличения содержания его в альвеолярном воздухе. В ре-зультате затрудняется выделение СО2 кровью, парциальное давление его в крови увеличивается, что вызывает большое возбуждение дыхательного центра и сильную одышку.
После задержки дыхания независимо от желания наступает временное усиление дыхания (гиперпноэ) вследствие накопления углекислоты в крови.
Уменьшение содержания углекислоты в крови после усиленного дыхания понижает возбудимость дыхательного центра и вызывает задержку дыхания (апноэ) до того момента, пока содержание углекислоты в крови не дойдет до обычного уровня.
Поэтому усиленное дыхание в течение 2 мин сопровождается последующей задержкой дыхания в течение 65—260 с. Апноэ наступает, когда содержание углекислого газа в альвеолярном воздухе понижается более чем на 0,2%. После спокойного дыхания человек может задержать дыхание приблизительно на 75 с. а тренированный человек значительно дольше.
Участие хеморецепторов в регуляции дыхания. Изменения напряжения кислорода и углекислоты в крови действуют на рецепторы кровеносных сосудов — хеморецепторы. Повышение напряжения углекислоты в крови (гиперкапния) раздражает хеморецепторы, расположенные в углу, образуемом разветвлением сонной артерии на наружную и внутреннюю, т. е. в каротидном синусе, что вызывает рефлекторное углубление дыхания. Понижение напряжения углекислоты крови (гипокапния), наоборот, вызывает с хеморецепторов каротидного синуса рефлекторное торможение дыхания. Недостаток кислорода в крови (гипоксемия), действуя на хеморецепторы каротидного синуса, приводит к учащению дыхания.
Асфиксия — удушье, характеризуется двумя фазами: недостатком кислорода в крови и избытком углекислоты в крови. В первой фазе наступает ннспираторная одышка, а во второй — экспираторная. Асфиксия первой фазы бывает при подъеме на большие высоты (на горы, в самолете). Асфиксия иногда наблюдается у плодов при нарушении мозгового кровообращения. Первый вдох новорожденного вызывается нарушением снабжения головного мозга кислородом после прекращения кровообращения через плаценту (перевязки пупочных сосудов).
Таким образом, дыхательный центр поддерживает на относительно постоянном уровне процентное содержание и парциальное давление углекислого газа в альвеолярном воздухе, а следовательно, и в крови. Это обеспечивает возбудимость нейронов самого дыхательного центра.
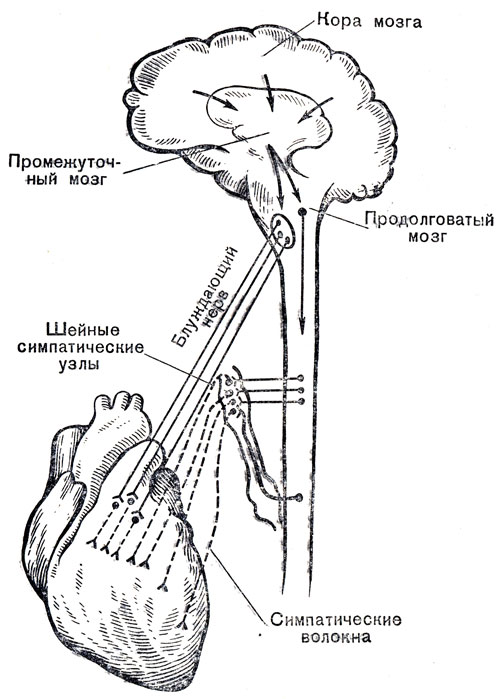
Особенности регуляции легочного дыхания у водных животных. У них имеются морфологические приспособления: резко увеличенная грудная клетка, особо мощная дыхательная мускулатура, большая подвижность грудной клетки, открывающиеся вверх носовые о тверстия (киты, дельфины). У дельфинов в 3 раза больше ильвсол, чем у человека. У китообразных в мелких бронхах есть мышечные жомы, длительно удерживающие воздух. Физиологические приспособления у водных животных следующие. Во-первых, Повышенная способность крови связывать кислород за счет увели-чения содержания гемоглобина в эритроцитах и увеличенной способности гемоглобина связывать кислород — например, у дельфинов и 1,5 раза. У них во много раз больше миоглобина, который и отличие от гемоглобина депонирует кислород и отдает его при необходимости: например, у тюленя содержание миоглобина н мышцах 20—40% от сухого остатка мышечной ткани. Во-вторых, при нырянии не только останавливается дыхание, но и резко уменьшается частота сердцебиений и происходит сужение кровеносных сосудов всего тела за исключением снабжающих кровью нервную систему, например у тюленя (рис. 15).
У всех ластоногих имеется специальный сфинктер из поперечнополосатой мышечной ткани, расположенный вокруг полой вены над диафрагмой. При нырянии он сдавливает полую вену и пре-кращает кровообращение во всем теле, кроме головы. Это обусловливает высокую устойчивость к большому содержанию) (Ж'кислоты в крови и сохраняет на прежнем уровне ее снабжение Кислородом. У ныряющих птиц также резко замедляются сердцебиения, они ритмически двигают под водой конечностями и крыльями, что вызывает перемешивание воздуха в воздушных мешках. В результате киты могут находиться под водой до 105, дельфины до 15, а утки до 23 мин
Цель исследования: оценить прогностическую роль последствий перинатальных нарушений центральной нервной системы (ППЦНС) у детей грудного возраста в формировании и особенностях течения аллергических заболеваний респираторного тракта. На базе поликлиники нами обследовано 42 ребенка от 2 до 12 мес. с ППЦНС (основная группа) и две группы сравнения, из которых 1-ю группу составили 27 детей с установленным диагнозом бронхиальной астмы легкой и средней степени тяжести в возрасте от 2,2 до 3 с половиной лет, 2-ю группу — 19 детей практически здоровых 1-2 А группы здоровья от 2 до 12 мес. без отягощенного перинатального и аллергологического анамнеза. Исследование проводилось по специально разработанной анкете. Для уточнения характера последствий перинатальной патологии нервной системы проводились: нейросонография (НСГ), компьютерная томография головного мозга, ЭЭГ, РЭГ, ПЭТ, УЗИ головного мозга. Отягощенность аллергоанамнеза оценивалась согласно критериям национальной программы «Бронхиальная астма у детей. Стратегия лечения и профилактики» (2008). Следует отметить, что у обследованных детей с отягощенным аллергоанамнезом данные факторы учитыва- лись в комплексе двух и более критериев. Результаты были обработаны при помощи пакета статистических программ Statistica 7.0. Все дети основной группы состояли на диспансерном учете у невролога с диагнозом: последствия ППЦНС легкой или средней тяжести. У всех детей основной группы и у 74% детей 1-й группы сравнения в анамнезе были диагностированы ППЦНС новорожденных: церебральная ишемия легкой и средней степени тяжести; внутричерепные кровоизлияния гипоксического генеза I степени (субэпендимальные). В качестве последствий ППЦНС у детей первого года жизни в основной и 1 группе сравнения наиболее часто встречались мышечные дистонии (гипо- и гипертонии как очаговые, так и генерализованные) (соответственно у 69,5% и 55% детей). Следует отметить, что в 1-й группе сравнения (с установленным диагнозом бронхиальной астмы) среди последствий ППЦНС наиболее часто (у 65% детей) наблюдался синдром вегетативных дисфункций. При сравнительной оценке патологических факторов течения беременности у детей основной и 1-й группы сравнения выявлена их идентичность (р>0,05). Патологическое течение родов (преждевременное излитие околоплодных вод, дискордантность родовой деятельности, обвитие пуповины вокруг шеи, быстрые роды, акушерские пособия, использование родостимуляции и др.) отмечалось в 85,7% случаев в основной группе и в 75,7% — в 1-й группе сравнения, что, безусловно, могло способствовать развитию перинатальных нарушений нервной системы. У большинства (83%) пациентов основной группы методом НСГ была диагностирована органическая патология головно- го мозга легкой и средней степени тяжести, нередко в виде сочетаний: нарушения оттока мозговой жидкости (конвекситальное скопление жидкости, нарушение венозного оттока, расширение желудочков) и кистозных образований (чаще в области желудочков и диэнцефально-гипоталамической области), нарушение церебрального кровообращения, расширение межполушарной борозды, повышенная резистентность сосудов головного мозга, вентрикуломегалия. В результате сопоставления уровней респираторной заболеваемости было выявлено, что дети основной и 1-й группы сравнения в 2 раза чаще болели ОРИ и в 12 раз чаще — бактериальными инфекциями на первом году жизни по сравнению с практически здоровыми детьми. Кроме того, нами отмечено, что у детей основной и 1-й группы сравнения коэффициент респираторной заболеваемости на первом году жизни (отношение количества ОРИ в течение первого года жизни к возрасту ребенка в мес.) был достоверно выше (соответственно 0,55±0,3 и 0,39±0,19), чем у практически здоровых детей (0,12±0,04; р
.
По данным исследований Мальцевой Е.А. и Михайлова Л.А. фактические значения показателей жизненной ёмкости лёгких, форсированной жизненной ёмкости лёгких, дыхательного объема у юношей – студентов достоверно выше, чем у девушек – студенток [28]. Выявлено снижение индивидуальных показателей жизненной ёмкости лёгких на вдохе и на выдохе, относительно должных величин у 60% и 37% студентов 18 – 20 лет, в результате исследования лёгочной вентиляции Силюк Ю.А. и Чудиновой О.В. Так же в ходе данного исследования установлено, что Ровд снижен, относительно должного у 67% юношей и девушек, а значение показателей ЧД, ДО и МОД преимущественно выше должных значений у студентов. Авторы связывают данные факты со слабым развитием мышечной системы, вследствие преобладания статических нагрузок.
Щербиной Ю.Ф. и Поповой О.Н. установлено у жителей Крайнего Севера, в юношеском возрасте, статистически значимое повышение минутной легочной вентиляции как у юношей (p < 0,001), так и у девушек (p = 0,042) в период полярной ночи по сравнению с периодом полярного дня [29]. Величина минутного объёма дыхания не является абсолютным показателем эффективности альвеолярной вентиляции, она является непосредственным результатом работы системы внешнего дыхания, которая обеспечивает поддержание скорости поступления кислорода в легкие и выведение углекислого газа на необходимом уровне. Легочная вентиляция находится в тесной зависимости от интенсивности процессов метаболизма в организме. Авторы считают, что для организма в юношеском возрасте, менее энергозатратным является механизм увеличения минутного объёма дыхания за счёт большего повышени