Кожная чувствительность
Рецепторы кожи
Кожные рецепторы являются первичными рецепторами и представляют собой окончания дендритов чувствительных нейронов, тела которых располагаются в спинномозговых ганглиях. Существует несколько классификаций рецепторов (нервных окончаний) кожи.
Гистологическая классификация основана на особенностях строения дендритов кожных рецепторов.
· Свободными нервными окончаниями называют дендриты чувствительных нейронов, лишенные глиальной капсулы.
· Окончания волосяных фолликулов представляет собой дендрит чувствительного нейрона, который спирально обкручивается вокруг основания волосяного фолликула.
· Инкапсулированные нервные окончания покрыты глиальной капсулой. В зависимости от устройства глиальной капсулы, различают
- тельца Мейснера,
- диски Меркеля,
- тельца Пачини,
- окончания Руффини.

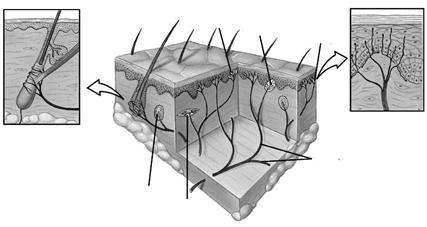
Рис. 4.1. Строение и расположение в коже разных типов рецепторов.
1 – дендрит чувствительного нейрона, 2 – свободное нервное окончание, 3 - тельце Мейснера, 4 - диски Меркеля, 5 – рецептор волосяного фолликула, 6 – тельце Пачини, 7 - окончание Руффини
Функциональная классификация делит рецепторы кожи в зависимости от воспринимаемого ими стимула.
· Механорецепторы воспринимают механическую деформацию кожи:
- медленно адаптирующиеся (рецепторы силы) реагируют на степень деформации кожи; частота генерируемых ими нервных импульсов возрастает пропорционально увеличению амплитуды деформации кожи (силе давления на кожу).
- быстро адаптирующиеся (рецепторы скорости) реагируют только на уменьшение или увеличения деформации кожи, постоянно действующее давление их не активирует; частота импульсации таких рецепторов пропорциональна скорости деформации кожи (скорости изменения давления на кожу)
- очень быстро адаптирующиеся (рецепторы ускорения) реагируют только на изменение скорости деформации кожи; как правило, они дают короткие залпы потенциалов действия в начале и в конце действия механического стимула.
· Терморецепторы воспринимают температуру кожи. Зависимость частоты импульсации терморецептора от температуры характеризуется куполообразной кривой:
- тепловые терморецепторы активируются при повышении температуры от 25 до 42 °С, дальнейшее увеличение температуры приводит к уменьшению частоты нервных импульсов;
- холодовые терморецепторы активируются при снижении температуры от 42 до 25 °С, дальнейшее охлаждение приводит к падению частоты нервных импульсов.
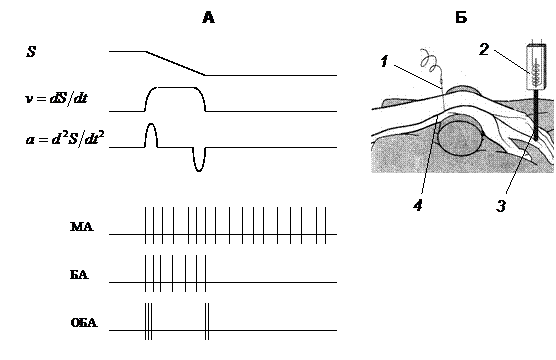
Рис. 4.2. Ответы четырех типов кожных механорецепторов на деформацию кожи (А) и схема установки для изучения кожной механической чувствительности кожи (Б).
A: три кривые в верхней части рисунка показывают динамику деформации кожи (S), скорость деформации (v) и ускорение деформации (а) в ходе одного цикла стимуляции; МА – медленно адаптирующиеся рецепторы (рецепторы силы), БА – быстро адаптирующиеся рецепторы (рецепторы скорости), ОБА - очень быстро адаптирующиеся рецепторы (рецепторы ускорения). Б: 1 – игольчатый электрод, введенный в нервное волокно лучевого нерва (4); 2 – электромеханостимулятор, который приводит в движение стержень (3), прогибающий кожу.
Проводящие пути
Аксоны чувствительных нейронов, иннервирующих кожу и мышцы туловища, входят в спинной мозг в составе задних корешков. Проведение возбуждения к коре больших полушарий (первичная и вторичная соматосенсорные проекционные зоны) осуществляется по двум восходящим системам:
· системе заднего столба (лемнисковая система),
· системе переднебокового канатика.
Сравнительная характеристика этих восходящих сенсорных путей представлена в табл. 3.
Таблица 3. Сравнительная характеристика проводящих путей соматосенсорной системы
| Система переднебокового канатика | Система заднего столба | |
| 1-е синаптическое переключение | Задние рога сегмента спинного мозга | Нежное и клиновидное ядро продолговатого мозга |
| 2-е синаптическое переключение | Вентробазальное ядро таламуса* | Вентробазальное ядро таламуса |
| 3-е синаптическое переключение | Кора больших полушарий* | Кора больших полушарий |
| Проводящие пути в составе спинного мозга | Спиноталамический тракт, спиноретикулярный тракт | Нежный пучок Голя, клиновидный пучок Бурдаха |
| Место перекреста проводящих путей | Сегмент спинного мозга | Ствол мозга выше нежного и клиновидного ядер (медиальный лемниск) |
| Конвергенция** | Мультимодальная | Мономодальная |
| Характер проводимой информации | Информация от терморецепторов кожи, болевых рецепторов | Информация от механорецепторов кожи, проприорецепторов |
| Скорость проведения | До 30 м/с | До 80 м/с |
* В системе переднебокового канатика информация к коре больших полушарий проводится по двум каналам (спиноталамический и спиноретикулярный). Спиноталамический путь организован по принципу соматотопической проекции и проходит через вентробазальный таламус. В таблице указаны синаптические переключения именно для этого пути. Спиноретикулярный путь проходит в составе ретикулярной формации ствола мозга, через медиальный таламус, образуя диффузные неспецифические проекции в коре больших полушарий. Этот путь является полисинаптическим, т.е. имеет большое количество переключений.
** Мультимодальная конвергенция подразумевает, что на одном и том же спинальном нейроне могут конвергировать афференты от рецепторов кожи, внутренних органов, мышечных рецепторов (см. рис. 4.5) При мономодальной конвергенции разные виды чувствительности не смешиваются – на одном спинальном нейроне заканчиваются афференты только одного вида рецепторов.
Обработка информации в ЦНС
Соматосенсорные функции ствола мозга. Кожа лица и шеи, слизистые рта и носа иннервируются 5-й парой черепно-мозговых нервов (тройничным нервом). На уровне ствола мозга афферентные волокна тройничного нерва вступают в ЦНС и направляются к ядрам тройничного нерва. Аксоны главного сенсорного ядра тройничного нерва образуют проекции к вентробазальному ядру таламуса, а нейроны сенсорного ядра тройничного нерва – в ретикулярную формацию и медиальный таламус.
Ретикулярная формация, в составе которой проходит часть восходящих путей переднебоковой системы, имеет множество физиологических функций, которые подробно обсуждаются в соответствующих лекциях.
 |
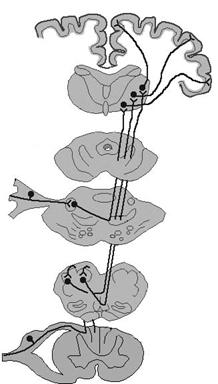
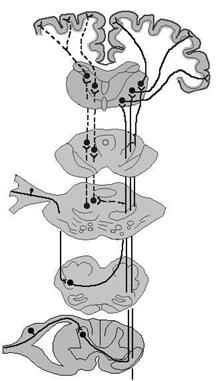
Рис. 4.3. Восходящие пути соматосенсорной системы.
Сплошными линиями показаны пути, организованные по принципу соматотопической проекции, пунктиром – диффузные проекции; МТ – медиальный таламус, ВБТ – вентробазальный таламус, РФ – ретикулярная формация, ТН – тройничный нерв, ЯТНГС – главное сенсорное ядро тройничного нерва, ЯТНС – спинальное ядро тройничного нерва.
Соматосенсорные функции таламуса. Через специфические ядра таламуса проходят все специфические восходящие сенсорные пути. Проекции в вентробазальном ядре таламуса организованы по соматотопическому принципу; каждый нейрон этого ядра имеет свое рецептивное поле на поверхности кожи. В специфических ядрах таламуса происходит начальная обработка всех видов сенсорной информации.
Соматосенсорные функции коры. Вентробазальное ядро таламуса образует строго упорядоченные проекции в первичную (SI) и вторичную (SII) соматосенсорную кору. В SI одного полушария поступает информация только с контрлатеральной стороны тела. В SII проекции билатеральны, т.е. в каждое полушарие поступают импульсы от рецепторов левой и правой половины тела.
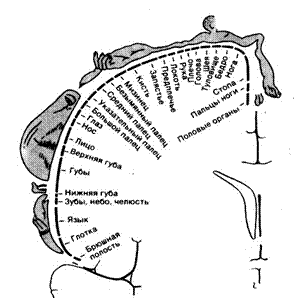
Рис 4.4. Соматотопическая организация корковой зоны SI человека. Изображения на поперечном срезе мозга и их обозначения демонстрируют пространственное представительство поверхности тела в коре.
Нейронные сети соматосенсорной коры выполняют две важные функции.
1. Выделение признаков сигнала. В зонах SI и SII обнаружены нейроны, которые реагируют на сложные характеристики стимулов, т.е. нейроны, которые узнают определенные воздействия. Например, описаны нейроны, которые возбуждаются только в ответ механические стимулы, перемещающиеся линейно о поверхности кожи и дают максимальный ответ на движение в определенном направлении.
2. Нисходящий контроль афферентного входа. Проведение информации по сенсорным путям находится под нисходящим контролем коры. Корковые нейроны образуют проекции в чувствительные ядра таламуса, нежное и клиновидные ядра ствола, задние рога спинного мозга. Эти нисходящие связи имеют преимущественно тормозный характер и могут вызывать следующие эффекты:
· повышение порога возбуждения нейронов проводящих путей, в результате этого происходит отфильтровывание фоновых раздражителей (например, мы не замечаем непрерывной стимуляции кожи одеждой);
· уменьшение рецептивных полей нейронов проводящих путей, что приводит к повышению четкости восприятия.
Проприорецепция
Типы проприорецепторов
У человека выделяют три типа проприорецепторов.
· Мышечные веретена представляют собой специализированные мышечные волокна, чувствительные к растяжению. Такие волокна окружены соединительнотканной капсулой, поэтому их называют интрафузальными. К каждому мышечному веретену подходит отросток спинального чувствительного нейрона (афферентное волокно группы Iа), который обвивается вокруг центральной части веретена. Мышечные веретена имеют также и двигательную иннервацию (т.н. альфа-гамма система, которая подробно рассматривается в лекции «Регуляция движений»).
· Сухожильные органы Гольджи состоят из сухожильных нитей, отходящих примерно от 10 экстрафузальных мышечных волокон и заключенных в соединительнотканную капсулу. К ней подходят 1-2 афферентных нервных волокна, относящихся к группе Ib. Войдя в капсулу, волокна образуют сильно разветвленные окончания среди сухожильных нитей.
· Суставные рецепторы располагаются в суставных сумках.
Электрические ответы проприорецепторов
Мышечные веретена в ненапряженной мышце генерируют нервные импульсы с некоторой частотой. При растяжении веретена частота нервных импульсов увеличивается, при уменьшении длины веретена (например, при сокращении мышцы) – уменьшается. Т.о. мышечные веретена воспринимают длину мышцы.
Сухожильные органы в состоянии покоя «молчат», они начинают генерировать импульсы при натяжении сухожилий, например, при сокращении мышцы или при ее пассивном растяжении под действием груза. Т.о. сухожильные органы воспринимают натяжение мышцы.
Суставные рецепторы активируются при изменении положения сустава.