Подготовили студентки 2 курса
Акинина Александра и Бедокурова Анастасия.
Фотодыхание. Основные аспекты в истории открытия. Отличия от темновой фазы.
Фотодыхание (гликолатный путь, С2-фотосинтез) — стимулируемое светом выделение углекислого газа и поглощение кислорода у растений преимущественно с С3-типом фотосинтеза. Также под фотодыханием понимают биохимический путь, связанный с регенерацией одной молекулы 3-фосфоглицериновой кислоты (С3) из двух молекул гликолевой кислоты (С2) и лежащий в основе вышеописанного газообмена. Наличие биохимического механизма фотодыхания обусловлено значительной оксигеназной активностью РуБисКО, ключевого фермента цикла Кальвина.
Поглощение кислорода в ходе фотодыхания обусловлено оксигеназной активностью РуБисКО в хлоропластах и работой оксидазы гликолевой кислоты в пероксисомах. Кроме того, окисление образовавшегося в митохондриях НАДН также сопряжено с поглощением кислорода. Выделение углекислого газа (С1) при фотодыхании происходит в митохондриях и связано с конденсацией двух молекул глицина (С2) с образованием одной молекулы серина (С3) (последовательная работа двух ферментов: глициндекарбоксилазы и серингидроксиметилтрансферазы). Также в реакции конденсации глицина в митохондриях выделяется аммиак, который реутилизируется в результате работы глутаминсинтетазы и глутаминоксоглутаратаминотрансферазы (ГС/ГОГАТ-путь). При фотодыхании расходуется АТФ (не происходит запасания энергии) синтезированный в ходе фотофосфорилирования. Также окисление гликолевой кислоты в пероксисомах в ходе фотодыхания служит основным источником токсичного пероксида водорода в фотосинтезирующей растительной клетке.
|
|
Первые свидетельства фотодыхания были получены в 1920 г. немецким биохимиком Отто Варбургом. При исследовании водорослей рода Chlorella было показано ингибирование фотосинтеза (поглощения углекислого газа) при повышении концентрации кислорода. Этот эффект наблюдался как при высоких, так и при низких интенсивностях света и позднее получил название эффекта Варбурга.
Явление фотодыхания открыто в 1955 г. И. Деккером. Оно в большей мере выражено у растений С3-пути фотосинтеза и сопровождается потерей углерода в цикле Кальвина из-за снижения способности РБФК фиксировать CO2 и повышения ее активности окислять субстрат (РБФ) в присутствии O2 (А. Лайск, 1977). В случае окисления субстрата РБФК выступает в роли РБФ-оксигеназы. На примере рода Panicum, в пределах которого встречаются виды с С3-, С4— и промежуточными путями фотосинтеза, выявлены различия в содержании органелл. У P. riculare (С3-путь) клетки обкладки содержат в среднем 3 хлоропласта, 1,1 митохондрий и 0,3 пероксисом; у Р. milioides, Р. decipiens и Р. schenckii (промежуточные луга) органелл соответственно 10,6,17,7 и 3,2; у Р. priontis (С4-путь) в клетках обкладки органелл не только больше, но они и крупнее. У указанных видов фотодыхание снижается в направлении увеличения числа органелл в клетках обкладки (R. Н. Brow et al., 1983).
Начальные стадии фотодыхания (образование гликолевой кислоты) характерны для всех организмов с циклом Кальвина. Виды же различаются механизмами метаболизма гликолевой кислоты. Так, фотосинтезирующие бактерии и цианеи окисляют гликолат и синтезируют глицин. Цианеи содержат фермент гликолатдегидрогеназу и способны поглощать гликолат (в темноте и на свету), но утилизируют его слабо. У эукариотических водорослей в процессе фотосинтеза гликолевая кислота выделяется в окружающую среду, хотя у них несколько лучше выражен механизм утилизации гликолата (при участии фермента гликолатдегидрогеназы). Подобное выделение представляет собой способ защиты цикла Кребса от ингибирования и на начальных этапах развития жизни было необходимо для усиления симбиотических отношений бактерий и цианей с гетеротрофами (А. М. Степанова, А. А. Шумилова, 1980).
|
|
У высших же растений с С3-путем фиксации CO2 фотодыхание осуществляется наиболее сложно — путем полного превращения гликолевой кислоты, с образованием ряда важных промежуточных продуктов (аминокислот, порфиринов и др.). Дело в том, что с переходом растений к наземным условиям существования возможности выделения гликолата в среду ограничиваются ввиду невымываемости его из организма. Поэтому защита цикла Кребса от ингибирующего действия глиоксилата могла быть достигнута только путем полного превращения его в гликолат.
Несмотря на эти новшества, у высших растений потери углерода при фотодыхании составляют 50% от его ассимилированного количества. У них фотосинтез падает главным образом из-за конкуренции O2 и CO2 за молекулу РБФК и ее субстрат. Интенсивность фотодыхания у растений С3-типа возрастает при повышении концентрации O2 и температуры среды. Это ведет к ингибированию фотосинтеза в результате снижения сродства РБФК к CO2, особенно в аридных условиях. Поэтому отбор способствовал формированию растений С4-типа, у которых хорошо развита система концентрации CO2 (ФЕП, органические кислоты и др.) и выражена активность РБФК (при одновременном подавлении оксигеназной функции указанного фермента). Благодаря подобным и другим изменениям у растений С4-типа потери углерода при фотосинтезе невелики и гликолат накапливается, медленно.
|
|
Хотелосьбы отметить, что ведущее место в фотодыхании принадлежит пероксисомам. Ни один другой органоид клетки не выполняет столь разнообразные функции, как микротельца (А. У. Игамбердиев, 1988, 1990), в том числе и при генерации новых путей биохимических превращений. Фотодыхание, хотя и рассматривается как ответвление фотосинтетического пути, играет важную роль в обеспечении нормального функционирования Цикла Кальвина, снабжая его CO2 в обход реакций карбоксилирования. Оно также устраняет ингибирование ЦТК метаболизацией гликолата. По этим и другим причинам необходимо было выработать механизм утилизации гликолата, от которого ранее (водоросли) избавлялись выделением в среду. С появлением пероксисом у части водорослей (хлорелла) окисление гликолата в глиоксилат с образованием H2O2 имело значение для рассеивания свободной энергии, поглощенной клеткой. Выделение гликолата в среду его вымыванием имело значение для развития симбиотических отношений автотрофов и гетеротрофов, но оно не могло получить дальнейшего развития в наземных условиях, где исключалась возможность этого вымывания. Отсюда и необходимость отбора растений в направлении усиления утилизации гликолата в энергетических целях. Косвенно эти механизмы способствовали усилению транспорта воды, минеральных и органических веществ у сосудистых растений благодаря формированию вакуолярной системы. Интересно отметить, что в случае повторного перехода цветковых к обитанию в воде они начинают выделять гликолат в среду.
Отличия от темнового дыхания.
Фотодыхание имеет ряд отличий от «темнового» дыхания клеток, связанного с процессами окисления органических соединений в ходе цикла Кребса и работы ЭТЦ митохондрий.
Во-первых, фотодыхание активируется светом высокой интенсивности, тогда как процессы «темнового» дыхания митохондрий на свету ингибируются.
Во-вторых, фотодыхание усиливается в присутствии высоких концентраций кислорода, что связано с активацией в этих условиях оксигеназной функции фермента РуБФ-карбоксилазы; максимум «темнового» дыхания наблюдается при концентрации кислорода около 2 %.
В-третьих, фотодыхание требует совместного функционирования хлоропластов, пероксисом и митохондрий; «темновое» дыхание связано исключительно с митохондриями.
В-четвертых, первичными продуктами фотодыхания являются фосфогликолат, гликолат и глиоксилат — двухуглеродные органические кислоты, дальнейшее преобразование которых приводит к образованию аминокислот глицина и серина.
Таким образом, суммарное поглощение кислорода на свету определяется интенсивностью двух процессов: темновым дыханием и светоиндуцируемым дыханием, связанным с фотохимическими реакциями хлоропластов. Фотодыхание активируется при высоких интенсивностях света, в то время как процессы темнового дыхания растений подавляются на свету. Фотодыхание увеличивается при повышении концентрации кислорода от 0 до 100 %, а темновое дыхание насыщается уже при 2 % кислорода. В отличие от темнового дыхания процесс поглощения кислорода при фотодыхании не ингибируется типичными дыхательными ядами, например азидом натрия (ингибитор цитохромоксидазы митохондрий).
Условия, стимулирующие фотодыхание
Очевидно, что снижение концентрации углекислого газа приводит к стимуляции фотодыхания. Как было сказано выше, фотодыхание интенсифицируется и при увеличении концентрации кислорода. Увеличение температуры приводит к снижению устойчивости ендиольного интермедиата реакции, катализируемой РуБисКО, что способствует реакции оксигенирования РуБФ. Помимо того, при повышении температуры растворимость углекислого газа уменьшается несколько сильнее растворимости кислорода (хотя и намного превышает её при любых температурах).
Субстратная специфичность рубиско
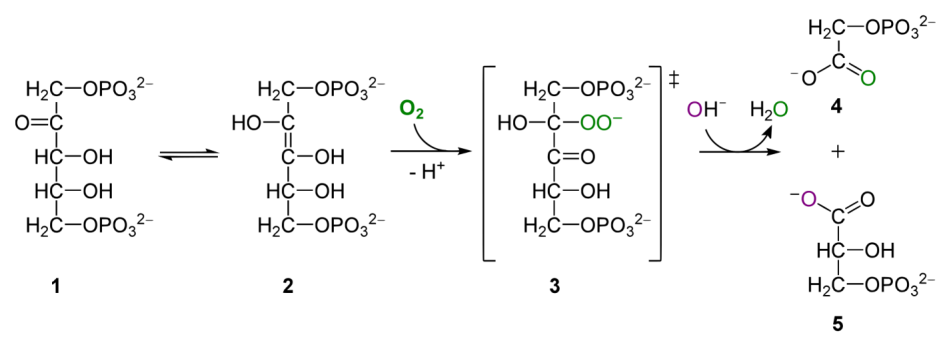
Рибулозобисфосфаткарбоксилаза/оксигеназа (Рубиско) — ключевой фермент фотосинтеза, катализирует две конкурирующие реакции: карбоксилирование и оксигенирование пятиуглеродного сахара рибулозо-1,5-бисфосфата (РуБФ). Молекула кислорода (как и углекислого газа) присоединяется к связанной с ферментом ендиольной форме РуБФ, которая существует вследствие кето-енольной изомерии. Константа Михаэлиса (концентрация полунасыщения) для углекислого газа гораздо ниже (9 мкМ), чем для кислорода (535 мкМ), то есть сродство фермента к углекислому газу значительно выше. Тем не менее скорость оксигеназной реакции высока, поскольку концентрация кислорода в атмосфере составляет 21 %, а углекислого газа — 0,04 %. Благодаря этому у С3-растений интенсивность фотодыхания может достигать 50 % от интенсивности фотосинтеза.
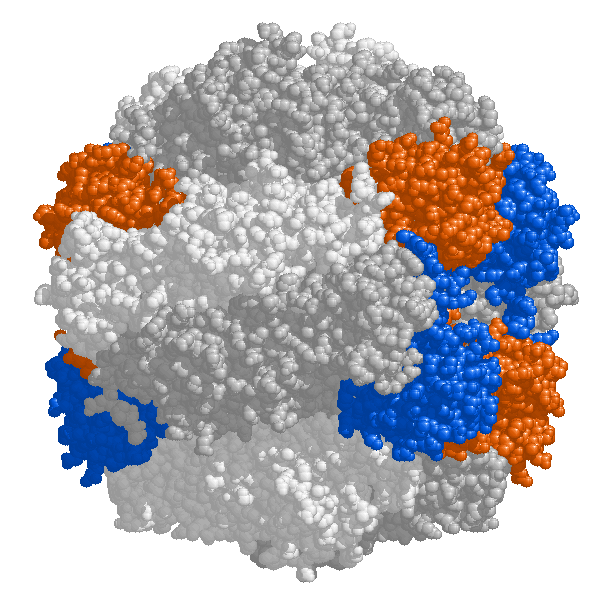 Цифровая модель фермента RuBisCO
Цифровая модель фермента RuBisCO
Структура:
У растений, цианобактерий и хемоавтотрофных протеобактерий фермент обычно состоит из двух типов белковых субъединиц:
- Большая цепь (L, массой 55 000 Да)
- Малая цепь (S, массой 13 000 Да)
Активный центр фермента находится на больших цепях, объединённых в димеры. Все 8 димеров больших цепей и 8 малых цепи объединены в единый комплекс массой 540 000 Да. У некоторых протеобактерий малых цепей в составе рубиско не обнаружено. При этом большие цепи кодируются в ДНК хлоропласта, а малые — в ядре и транспортируются в хлоропласт перед сборкой всего белка.
Для работы фермента необходимы ионы Mg2+, которые размещаются в активном центре и способствуют присоединению CO2 к остатку лизина, в ходе чего образуется карбамат. Образование карбамата протекает легче в щелочной среде: роль pH и ионов магния в регуляции работы фермента описано ниже.
Каталитическое действие:
Субстратами для рибулозобифосфаткарбоксилазы являются рибулозо-1,5-бисфосфат, углекислый газ и вода, вместо углекислого газа может быть метаболизирован молекулярный кислород.
- В случае углекислого газа первым промежуточным продуктом выступает нестабильный шестиуглеродный фосфорилированный сахар, который сразу же распадается на две молекулы 3-фосфоглицериновой кислоты (3-фосфоглицерата). Фосфоглицерат используется для синтеза глюкозы.
- В случае кислорода продуктами реакции являются 3-фосфоглицерат и фосфат гликолевой кислоты (ФГК). ФГК растения далее используют в процессе фотодыхания.
Реакция, проводимая ферментом, протекает относительно медленно (используются лишь несколько молекул углекислого газа в секунду) и является лимитирующей стадией всего цикла Кальвина. Константа Михаэлиса для карбоксилазной реакции рибулозобисфосфаткарбоксилазы равна 10±4 мкМ CO2, для оксигеназной 0,5 мМ O2, для рибулозобифосфата 1,5±0,5 мкМ.
Недостатки:
1. Рубиско нетороплив — каждую секунду этот фермент обрабатывает лишь от 3 до 10 молекул углекислого газа, в зависимости от условий. Понятно, что такое низкое качество работы фермента можно компенсировать лишь его количеством, и потому хлоропласты под завязку забиты этим белком: по массе на него приходится до 30% всех водорастворимых белков растений, что делает его самым распространенным белком на планете.
2. Крайне неразборчив и частенько путает углекислый газ с кислородом. И тогда вместо присоединения атома углерода к фосфорилированному сахару с наращиванием его углеродной цепи он просто окисляет этот сахар (рибулоза-(1,5)-бифосфат), попусту сжигая ценные молекулы углевода. В результате этого процесса, называемого фотодыханием, часть энергии, запасенной во время фотосинтеза, расходуется, причем потери КПД иногда достигают аж 25%, т.е. каждый четвертый углерод, связанный из СО2 при фотосинтезе, затем опять окисляется из-за фотодыхания. Особенно драматично эффект проявляется при высокой температуре, так что некоторым растениям пришлось даже оптимизировать строение своих листьев и включить в фотосинтез дополнительные реакции, призванные поднять в хлоропластах концентрацию углекислого газа и снизить концентрацию кислорода, тем самым увеличив эффективность неповоротливого фермента.
3. Получается, что работа Рубиско становится лимитирующей стадией всего фотосинтеза.