С.А. Апрятин. Основные принципы генетики и селекции аквариумных рыб.
Основные принципы генетики и селекции аквариумных рыб.
К.б.н. С.А. Апрятин
(журнал "Аквариум".- №3.- 2010)
На написание этой статьи меня «подбили» многочисленные обращения знакомых и незнакомых аквариумистов, которые очень хотели бы заниматься селекцией аквариумных рыб, но для этого не хватало теоретических знаний и практического опыта. Таких людей оказалось достаточно много, и разбирать подробно с каждым все премудрости генетики и селекции просто не представлялось возможным. И если практический опыт приходит с годами, то теоретические знания можно получить в любое время. Как говорится, было бы желание. В этой статье мне хотелось бы кратко коснуться основных понятий общей генетики и селекции, а также поговорить о генетическом анализе – методе, без которого серьезная селекция превращается в простое наблюдение и отбор особей по внешнему виду (фенотипу) без понимания генетики, которая заключается, прежде всего, в определении количества и типа взаимодействия генов, отвечающих за развитие тех или иных признаков. Для начала необходимо освоить теоретические азы генетики и селекции. Можно читать разные книги, просматривать бесконечные интернет-страницы, можно общаться на форумах, знакомиться с опытными селекционерами и т.д. Вариантов может быть много, и выбор предоставлен вам. Далее определяемся с тем, что вы хотите сделать? Например, получить новую селекционную линию с определенными свойствами (размер и форма плавников, цвет, пятна, сложный узор и т.п.). После этого необходимо найти подходящих производителей. Делать это желательно не в зоомагазине или на рынке (вариант "купил у дяди"), а у проверенных опытных разводчиков, хорошо знающих свою рыбу. Но даже в случае покупки рыбы с неизвестной родословной на помощь придет знание генетического анализа. Просто на определение генотипа рыб (а значит и на селекционную работу) вы потратите больше времени. Важно понимать, что каждая селекционная линия и каждая порода уникальны в своем роде. Нельзя проводить селекцию любых пород и селекционных линий по одним и тем же критериям. Каждый раз необходимо начинать с нуля, анализируя потомство, полученное от каждых производителей в первом и втором поколениях с помощью генетического анализа, с основными принципами которого вы познакомитесь в этой статье. Но прежде чем перейти к самому генетическому анализу, необходимо знать и понимать основные термины, закономерности и правила общей генетики. Для начала определимся с основными понятиями в генетике и селекции. Что же такое ген? Ген – это структурная единица наследственности, фрагмент ДНК (двухцепочный полимер, который в комплексе с белками образует хромосомы), контролирующий развитие того или иного признака - фена. Каждый ген занимает определенное место в хромосоме. Фактически в гене закодирована информация о белке (ферменте, переносчике, рецепторе, гормоне (если он имеет белковую природу), структурном элементе и т.п.), имеющем определенную функцию в организме. Гены обозначают либо прописной буквой латинского алфавита - A, B, C и т.д., либо сокращенно 2-3 буквами от названия гена, например, ген Ma (от лат. "Maculatus” – пятнистый, крапчатый). Подавляющее большинство генов "скрыты" в хромосомах ядра клетки (некоторые гены находятся в специальных клеточных органеллах – митохондриях, которые отвечают за процесс выработки клеткой энергии). Мы можем судить о генах только по проявлению признаков и свойств, которые ими и определяются. Эти признаки представляют собой не только общепринятые свойства организма, такие как цвет, узор форма, размер. Среди них можно выделить плодовитость, иммунитет, особенности поведения (агрессивность, половая активность, забота о потомстве и т.п.), устойчивость к изменениям температуры воды, стрессам и многие другие. Таким образом, проявляющийся признак называют доминантным, а не проявляющийся (скрытый) – рецессивным. А такие же варианты одного и того же гена – доминантным и рецессивным аллелями, соответственно. Аллелей (вариантов) у каждого гена может быть два и больше. Исключение составляют только половые хромосомы (о генетике которых мы еще поговорим в этой статье), которые могут содержать как один, так и два (или более) аллеля одного гена. Доминантный аллель гена обозначается обычно заглавной буквой (А), а рецессивный – строчной (а). Если аллелей несколько (множественный аллелизм), то они обозначаются соответствующей буквой с числовым индексом – А1, А2, а1, а2 и т.д.. Несколько слов нужно сказать и об условных обозначениях, применяемых в селекционных схемах. Родители обозначаются буквой P (от лат. "Parental”), дети – F (от лат. "Filli”), F1, F2 и т.д. – первое, второе поколения и т.д. Если в схеме не указан пол особей, то на первом месте в скрещивании стоит самка. Как уже говорилось выше, гены находятся в хромосомах. У разных видов их количество отличается. Интересно, что существует тенденция увеличения количества хромосом морских и особенно пресноводных рыб от экватора к полюсам. По всей видимости, это связано с большим генетическим разнообразием, которое жизненно необходимо в суровых условиях высоких широт. Все хромосомы делятся на аутосомы и гоносомы (половые хромосомы – X и Y). Все они сходны по строению, однако половые хромосомы, в отличие от аутосом, имеют как общие, так и различные гены (которые присутствуют только в одной из половых хромосом). Если наследование признака осуществляется независимо от того, какой из родителей является носителем изучаемого признака, то признак считается аутосомным, а если наоборот, то частично (XY-сцепленность) или полностью (X- или Y-сцепленность) сцепленным с полом (половыми хромосомами). В последнем случае наследование признака у самцов и самок будет различным. Важно отметить, что у разных видов рыб пол определяется по-разному: в одном случае самец несет как X-, так и Y-хромосому, а самка две X-хромосомы, в другом - наоборот (при этом хромосомы называются Z и W, но принципиальных отличий от X- и Y-хромосом нет). Как известно, жизнь многоклеточного организма начинается с одной оплодотворенной яйцеклетки – зиготы, деление которой обеспечивает рост и развитие всего организма. В основе такого деления лежит митоз, при котором количество клеток удваивается, при этом количество генетической информации в каждой клетке остается неизменным – они несут по одному полному набору хромосом и от отца и от матери - такой набор называется диплоидным (двойной набор) и обозначается «2n» и характерен для клеток всего организма (рис. 1).
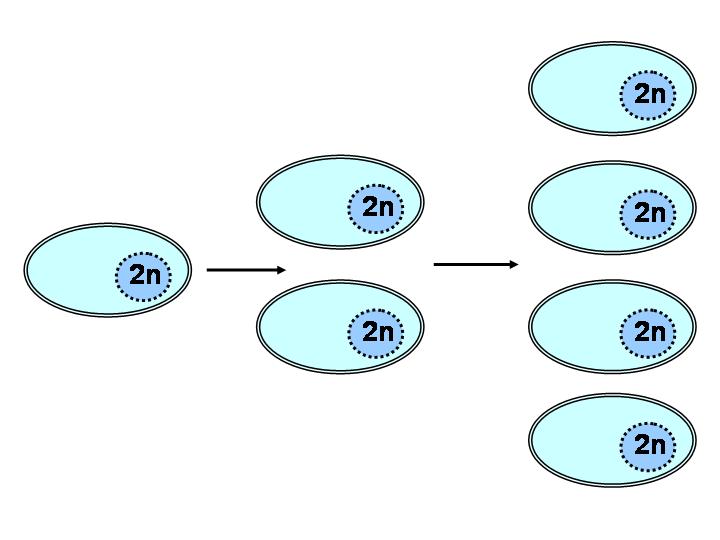 Рис.1. Митоз.
В отличие от митоза в процессе мейоза (образования половых клеток – гамет) количество генетического материала в дочерних клетках уменьшится вдвое – становится гаплоидным (одиночный набор) и обозначается буквой «n» (рис.2). При половом размножении происходит слияние половых клеток (гамет) с гаплоидным числом хромосом и восстанавливается диплоидное количество (рис.3).
Рис.1. Митоз.
В отличие от митоза в процессе мейоза (образования половых клеток – гамет) количество генетического материала в дочерних клетках уменьшится вдвое – становится гаплоидным (одиночный набор) и обозначается буквой «n» (рис.2). При половом размножении происходит слияние половых клеток (гамет) с гаплоидным числом хромосом и восстанавливается диплоидное количество (рис.3).
 Рис. 2. Мейоз.
Рис. 2. Мейоз.
 Рис.3. Образование зиготы.
То, что многие признаки передаются по наследству, было известно тысячи лет назад, однако только в 1866 году Грегором Иоганном Менделем были открыты законы наследственности. Интересно, что его работа оставалась незамеченной более 30 лет, когда его законы и правило были заново открыты сразу тремя учеными. Вот эти законы:
1-й закон Менделя– закон единообразия гибридов первого поколения. При скрещивании особей, различающихся по одной паре признаков, за которые отвечают аллели одного гена, первое поколение гибридов одинаково по фенотипу и генотипу. Таким образом, все гибриды первого поколения имеют доминантный фенотип и гетерозиготный генотип.
2-й закон Менделя– закон расщепления гибридов второго поколения. При скрещивании особей, различающихся по одной паре признаков, за которые отвечают аллели одного гена, во втором поколении наблюдается расщепление по фенотипу в соотношении 3:1. При этом около ¾ гибридов имеют доминантный признак, а ¼ - рецессивный.
3-й закон Менделя– закон независимого наследования (комбинирования) признаков. Каждая пара признаков наследуется независимо от других пар и дает расщепление 3:1 по каждой паре. Таким образом, для одного гена общее расщепление будет 3:1, для двух – 9:3:3:1, для трех – 27:9:9:9:3:3:3:1 и т.д. (таблица 1). Также существует правило чистоты гамет. Оно гласит: в каждую пару гамет (половых клеток) попадает только один аллель из пары аллелей одного гена (от каждого из родителей).
Рис.3. Образование зиготы.
То, что многие признаки передаются по наследству, было известно тысячи лет назад, однако только в 1866 году Грегором Иоганном Менделем были открыты законы наследственности. Интересно, что его работа оставалась незамеченной более 30 лет, когда его законы и правило были заново открыты сразу тремя учеными. Вот эти законы:
1-й закон Менделя– закон единообразия гибридов первого поколения. При скрещивании особей, различающихся по одной паре признаков, за которые отвечают аллели одного гена, первое поколение гибридов одинаково по фенотипу и генотипу. Таким образом, все гибриды первого поколения имеют доминантный фенотип и гетерозиготный генотип.
2-й закон Менделя– закон расщепления гибридов второго поколения. При скрещивании особей, различающихся по одной паре признаков, за которые отвечают аллели одного гена, во втором поколении наблюдается расщепление по фенотипу в соотношении 3:1. При этом около ¾ гибридов имеют доминантный признак, а ¼ - рецессивный.
3-й закон Менделя– закон независимого наследования (комбинирования) признаков. Каждая пара признаков наследуется независимо от других пар и дает расщепление 3:1 по каждой паре. Таким образом, для одного гена общее расщепление будет 3:1, для двух – 9:3:3:1, для трех – 27:9:9:9:3:3:3:1 и т.д. (таблица 1). Также существует правило чистоты гамет. Оно гласит: в каждую пару гамет (половых клеток) попадает только один аллель из пары аллелей одного гена (от каждого из родителей).
 Таблица 1. Число образующихся типов и комбинаций гамет, а также фенотипов и генотипов при различных типах скрещиваний.
Теперь, когда мы знаем, что гены (и их аллели) взаимодействуют между собой и каковы общие принципы этих взаимодействий, перейдем на следующий уровень в понимании этого вопроса. Взаимодействие бывает аллельным и неаллельным. Аллельное взаимодействие делится на полное доминирование (один аллель полностью доминирует над другим аллелем), неполное доминирование (доминантный аллель не полностью скрывает рецессивный, и признаки частично смешиваются, давая промежуточный фенотип) и кодоминирование (оба аллеля в одинаковой мере проявляются в фенотипе). Напомню, что в случае аллельного взаимодействия мы говорим о наследовании аллелей одного гена. Неаллельное взаимодействие имеет совсем другой характер. Здесь нет доминирования в классическом понимании этого термина, а различные типы взаимодействий идут между аллелями разных генов. Среди них можно выделить эпистаз, комплементарность и полимерию. Несколько слов также стоит сказать о наследовании генов, сцепленных с полом. Но обо всем по порядку.
Эпистаз представляет собой взаимодействие двух неаллельных (разных) генов. Один аллель гена подавляет экспрессию другого аллеля таким образом, что кодируемый им признак либо утрачивается, либо изменяется. Фенотипически эпистаз выражается в отклонении от ожидаемого расщепления, однако нарушение законов Менделя в этом случае не происходит, поскольку распределение аллелей взаимодействующих генов полностью соответствует закону независимого комбинирования. Различают следующие виды эпистаза: простой рецессивный эпистаз - a>B, a>b, расщепление 9:3:4 двойной рецессивный эпистаз - a>B, a>b, b>A, b>a, расщепление 9:7 простой доминантный эпистаз - A>B, A>b, расщепление 12:3:1 Знаком «>» показано подавление действия одного аллеля другим.
Комплементарность – это тип взаимодействия между генами, при котором доминантные аллели разных генов, взаимно дополняя друг друга, обуславливают определенный признак (фенотип).
Полимерия - взаимодействие двух неаллельных (разных) генов, при котором оба из них дублируют действие друг друга, и одного из доминантных аллелей любого из взаимодействующих генов достаточно для проявления изучаемого признака. Различают кумулятивную (когда количество однозначно действующих доминантных аллелей взаимодействующих генов в генотипе будет (пропорционально) определять степень развития изучаемого признака) и некумулятивную (когда степень развития признака определяется наличием в генотипе хотя бы одного доминантного аллеля любого из взаимодействующих генов) полимериии. Важно отметить, что наследование по законам Менделя происходит когда изучаемые гены находятся в разных хромосомах. В этом случае работает закон независимого комбинирования признаков. Но если гены расположены в одной хромосоме, то они наследуются вместе (сцеплено) и независимого комбинирования признаков происходить не будет. Гены окажутся связанными в тех сочетаниях, в каких они были у исходных родителей. В мейозе существует процесс, который необходим для повышения степени генетического разнообразия организма – называется он кроссинговер и его изучение является темой для отдельной статьи. Таким образом, общим выводом вышесказанного является понимание того, что селекционер должен анализировать пару признаков (доминантный и рецессивный) для каждого изучаемого гена, а не один (интересующий его) из них. Именно это является одной из частых ошибок начинающего селекционера.
Теперь перейдем к изучению генетического анализа, который является методологической основой генетики. Известный русский генетик А.С. Серебровский писал: "Лучшим способом овладения методами генетического анализа является экспериментальная работа с модельными объектами и решение генетических задач".С этими словами трудно не согласиться, также как и с другим его тезисом: "Генетические задачи решаются легко только тогда, когда они предварительно уже решены другими. Поэтому необходимо предостеречь тех, кто впервые приступает к генетическому анализу, от уныния и пессимизма, если их первые опыты окажутся неудачными".Поэтому в этой статье предлагаю вам, уважаемые аквариумисты-селекционеры, познакомиться с основами генетики и селекции, освоить азы генетического анализа, что является необходимым условием для успешной селекции аквариумных рыб (и любых других животных и растений). Генетический анализ подразумевает, прежде всего, оценку первого и второго поколений, а также анализ различных вспомогательных скрещиваний (анализирующего, возвратного и др.). Важно отметить, что на основе сходства первого поколения с одним из родителей еще нельзя говорить о доминантности признака. Мендель действительно предложил термин "доминантность" для обозначения признака, проявляющегося в первом поколении (F1). Но при этом подразумевалось, что каждый признак находится под контролем одного гена. Уже после открытий Менделя был открыт полигенный контроль большинства признаков, а термин "доминантный" сейчас применяется только для обозначения аллельных взаимодействий (например, «А» и «а», но не «В» и «а»). Если генов 2 и больше - то потомство F1 может также быть похожим на одного из родителей, но это сходство является следствием разных типов неаллельных взаимодействий, например, доминантного эпистаза. Принцип генетического анализа состоит в получении наследственно различающихся по определенным признакам форм и изучение этих различий. Основная задача генетического анализа – изучение наследования отдельных признаков для установления гена. Итак, например, вы определились с теми признаками, которые хотите получить. Для того чтобы грамотно провести генетический анализ необходимо выполнение ряда обязательных условий:
1. Подбор производителей.
2. Генетический анализ первого поколения (F1).
3. Генетический анализ второго поколения (F2).
4. Постановка дополнительных скрещиваний (если это необходимо).
5. Итоговый генетический анализ полученных результатов.
Разберем все условия более подробно.
1. Подбор производителей.
Для классического генетического анализа необходимо выбирать гомозиготных по исследуемым признакам родителей, т.е. необходимо брать чистопородных рыб. Рыбы должны быть взрослыми, здоровыми, без видимых патологий. Для селекции не следует ограничиваться одной парой производителей. Для дальнейшего генетического анализа лучше взять несколько пар производителей, поскольку при полигенном наследовании изучаемых признаков (3 и более генов) кол-во возможных классов в F1 и, особенно, в F2 может быть настолько большим, что потомство из 20-40 особей (например, у живородящих) будет содержать только малую их часть, в то время как большая часть этих классов просто не будет представлена в потомстве. Казалось бы, это не должно относится ко многим икромечущим аквариумным рыбам (американским и африканским цихлидам, лабиринтовым и др.), дающим сотни и тысячи икринок в одном нересте. Однако не стоит забывать, что генетическое разнообразие является необходимым условием для создания новой породы или селекционной линии. Таким образом, в зависимости от вида рыб, репродуктивных и других индивидуальных особенностей производителей селекционер выбирает оптимальное количество пар и таким образом закладывает фундамент для будущего генофонда селекционной линии.
2. Анализ первого поколения (F1).
Итак, мы отобрали и скрестили производителей, получили и вырастили потомство. Для начала необходимо посчитать общее количество полученных рыб в F1, а также отдельно количество самцов и самок. Далее нужно подсчитать общее количество рыб с каждым из исследуемых признаков, а также отдельно по самцам и самкам. Если отличий между общим количеством рыб и отдельно самками и самцами нет, скорее всего, исследуемый признак аутосомный. Если есть отличие в расщеплении по самцам и самкам, то признак полностью или частично сцеплен с полом. После этого необходимо скрестить братьев и сестер (F1) между собой. Как уже было сказано выше, необходимо взять несколько пар производителей (для живородящих 5-6 пар и более), получить и вырастить второе поколение и приступить к генетическому анализу F2.
3. Анализ второго поколения (F2).
В первую очередь необходимо понять какое расщепление произошло в F2. Как многим могло показаться, что ничего сложного здесь нет. Например, от креста F1 (родители имели зеленый и желтый окрас) во втором поколении было получено 26 мальков, среди которых наблюдалось следующее расщепление: 14 синих, 10 зеленых и 2 желтых. Что сначала приходит в голову? 7:5:1. Но это неправильный ход рассуждений. Почему? Потому что для подсчета соотношений в F2 существуют определенные правила. Для начала необходимо предположить количество генов, участвующих в определении признака. Как это сделать? Для этого нужно посмотреть, сколько фенотипических классов образовалось в F2 и каково их примерное соотношение. Если образовалось более 3-х классов, то можно с уверенностью говорить, что в определении признака участвует 2 и более генов. Однако наличие 2-3 классов в F2 также не всегда говорит о том, что это разные аллельные варианты одного гена (конечно, если это не 3:1 или 1:2:1). Как правильно посчитать соотношение? Сначала нужно определить (предположить) примерное количество генов, участвующих в определении признака. Далее нужно подсчитать количество возможных вариантов генотипов, которые образуются в F2. Это делается по формуле: Х= 4 в степени "n" (таблица 1), где n - предполагаемое количество генов, а Х - количество возможных вариантов генотипов. В нашем примере мы предполагаем, что в наследовании признака окраса участвуют 2 гена, значит Х=16. Действительно, по количеству мальков в трех фенотипических классах (синие, красные и желтые) расщепление отличается от неполного доминирования (1:2:1). Общее кол-во рыб в F2 (а также отдельное количество самок и самцов) делится на 16 (14+10+2 / 16) и мы получаем 1/16 часть = 1,6. Учитывая, что возможных вариантов генотипов может быть 16, мы должны разделить количество рыб в каждом классе на 1,6: синие - 14/1,6=8,7; зеленые - 10/1,6=6,2; желтые - 2/1,6=1,2, что примерно равно отношению 9:6:1. Мы получили истинное расщепление 9:6:1, а не 7:5:1. Возможное сочетания гамет и генотипов поможет выявить так называемая решетка Пеннета, используя которую можно легко понять взаимодействие аллелей двух генов. По вертикали в ней расписаны гаметы самца, а по горизонтали – самки. Таким образом, в клетках решетки показаны все возможные варианты генотипов (рис. 4).
Таблица 1. Число образующихся типов и комбинаций гамет, а также фенотипов и генотипов при различных типах скрещиваний.
Теперь, когда мы знаем, что гены (и их аллели) взаимодействуют между собой и каковы общие принципы этих взаимодействий, перейдем на следующий уровень в понимании этого вопроса. Взаимодействие бывает аллельным и неаллельным. Аллельное взаимодействие делится на полное доминирование (один аллель полностью доминирует над другим аллелем), неполное доминирование (доминантный аллель не полностью скрывает рецессивный, и признаки частично смешиваются, давая промежуточный фенотип) и кодоминирование (оба аллеля в одинаковой мере проявляются в фенотипе). Напомню, что в случае аллельного взаимодействия мы говорим о наследовании аллелей одного гена. Неаллельное взаимодействие имеет совсем другой характер. Здесь нет доминирования в классическом понимании этого термина, а различные типы взаимодействий идут между аллелями разных генов. Среди них можно выделить эпистаз, комплементарность и полимерию. Несколько слов также стоит сказать о наследовании генов, сцепленных с полом. Но обо всем по порядку.
Эпистаз представляет собой взаимодействие двух неаллельных (разных) генов. Один аллель гена подавляет экспрессию другого аллеля таким образом, что кодируемый им признак либо утрачивается, либо изменяется. Фенотипически эпистаз выражается в отклонении от ожидаемого расщепления, однако нарушение законов Менделя в этом случае не происходит, поскольку распределение аллелей взаимодействующих генов полностью соответствует закону независимого комбинирования. Различают следующие виды эпистаза: простой рецессивный эпистаз - a>B, a>b, расщепление 9:3:4 двойной рецессивный эпистаз - a>B, a>b, b>A, b>a, расщепление 9:7 простой доминантный эпистаз - A>B, A>b, расщепление 12:3:1 Знаком «>» показано подавление действия одного аллеля другим.
Комплементарность – это тип взаимодействия между генами, при котором доминантные аллели разных генов, взаимно дополняя друг друга, обуславливают определенный признак (фенотип).
Полимерия - взаимодействие двух неаллельных (разных) генов, при котором оба из них дублируют действие друг друга, и одного из доминантных аллелей любого из взаимодействующих генов достаточно для проявления изучаемого признака. Различают кумулятивную (когда количество однозначно действующих доминантных аллелей взаимодействующих генов в генотипе будет (пропорционально) определять степень развития изучаемого признака) и некумулятивную (когда степень развития признака определяется наличием в генотипе хотя бы одного доминантного аллеля любого из взаимодействующих генов) полимериии. Важно отметить, что наследование по законам Менделя происходит когда изучаемые гены находятся в разных хромосомах. В этом случае работает закон независимого комбинирования признаков. Но если гены расположены в одной хромосоме, то они наследуются вместе (сцеплено) и независимого комбинирования признаков происходить не будет. Гены окажутся связанными в тех сочетаниях, в каких они были у исходных родителей. В мейозе существует процесс, который необходим для повышения степени генетического разнообразия организма – называется он кроссинговер и его изучение является темой для отдельной статьи. Таким образом, общим выводом вышесказанного является понимание того, что селекционер должен анализировать пару признаков (доминантный и рецессивный) для каждого изучаемого гена, а не один (интересующий его) из них. Именно это является одной из частых ошибок начинающего селекционера.
Теперь перейдем к изучению генетического анализа, который является методологической основой генетики. Известный русский генетик А.С. Серебровский писал: "Лучшим способом овладения методами генетического анализа является экспериментальная работа с модельными объектами и решение генетических задач".С этими словами трудно не согласиться, также как и с другим его тезисом: "Генетические задачи решаются легко только тогда, когда они предварительно уже решены другими. Поэтому необходимо предостеречь тех, кто впервые приступает к генетическому анализу, от уныния и пессимизма, если их первые опыты окажутся неудачными".Поэтому в этой статье предлагаю вам, уважаемые аквариумисты-селекционеры, познакомиться с основами генетики и селекции, освоить азы генетического анализа, что является необходимым условием для успешной селекции аквариумных рыб (и любых других животных и растений). Генетический анализ подразумевает, прежде всего, оценку первого и второго поколений, а также анализ различных вспомогательных скрещиваний (анализирующего, возвратного и др.). Важно отметить, что на основе сходства первого поколения с одним из родителей еще нельзя говорить о доминантности признака. Мендель действительно предложил термин "доминантность" для обозначения признака, проявляющегося в первом поколении (F1). Но при этом подразумевалось, что каждый признак находится под контролем одного гена. Уже после открытий Менделя был открыт полигенный контроль большинства признаков, а термин "доминантный" сейчас применяется только для обозначения аллельных взаимодействий (например, «А» и «а», но не «В» и «а»). Если генов 2 и больше - то потомство F1 может также быть похожим на одного из родителей, но это сходство является следствием разных типов неаллельных взаимодействий, например, доминантного эпистаза. Принцип генетического анализа состоит в получении наследственно различающихся по определенным признакам форм и изучение этих различий. Основная задача генетического анализа – изучение наследования отдельных признаков для установления гена. Итак, например, вы определились с теми признаками, которые хотите получить. Для того чтобы грамотно провести генетический анализ необходимо выполнение ряда обязательных условий:
1. Подбор производителей.
2. Генетический анализ первого поколения (F1).
3. Генетический анализ второго поколения (F2).
4. Постановка дополнительных скрещиваний (если это необходимо).
5. Итоговый генетический анализ полученных результатов.
Разберем все условия более подробно.
1. Подбор производителей.
Для классического генетического анализа необходимо выбирать гомозиготных по исследуемым признакам родителей, т.е. необходимо брать чистопородных рыб. Рыбы должны быть взрослыми, здоровыми, без видимых патологий. Для селекции не следует ограничиваться одной парой производителей. Для дальнейшего генетического анализа лучше взять несколько пар производителей, поскольку при полигенном наследовании изучаемых признаков (3 и более генов) кол-во возможных классов в F1 и, особенно, в F2 может быть настолько большим, что потомство из 20-40 особей (например, у живородящих) будет содержать только малую их часть, в то время как большая часть этих классов просто не будет представлена в потомстве. Казалось бы, это не должно относится ко многим икромечущим аквариумным рыбам (американским и африканским цихлидам, лабиринтовым и др.), дающим сотни и тысячи икринок в одном нересте. Однако не стоит забывать, что генетическое разнообразие является необходимым условием для создания новой породы или селекционной линии. Таким образом, в зависимости от вида рыб, репродуктивных и других индивидуальных особенностей производителей селекционер выбирает оптимальное количество пар и таким образом закладывает фундамент для будущего генофонда селекционной линии.
2. Анализ первого поколения (F1).
Итак, мы отобрали и скрестили производителей, получили и вырастили потомство. Для начала необходимо посчитать общее количество полученных рыб в F1, а также отдельно количество самцов и самок. Далее нужно подсчитать общее количество рыб с каждым из исследуемых признаков, а также отдельно по самцам и самкам. Если отличий между общим количеством рыб и отдельно самками и самцами нет, скорее всего, исследуемый признак аутосомный. Если есть отличие в расщеплении по самцам и самкам, то признак полностью или частично сцеплен с полом. После этого необходимо скрестить братьев и сестер (F1) между собой. Как уже было сказано выше, необходимо взять несколько пар производителей (для живородящих 5-6 пар и более), получить и вырастить второе поколение и приступить к генетическому анализу F2.
3. Анализ второго поколения (F2).
В первую очередь необходимо понять какое расщепление произошло в F2. Как многим могло показаться, что ничего сложного здесь нет. Например, от креста F1 (родители имели зеленый и желтый окрас) во втором поколении было получено 26 мальков, среди которых наблюдалось следующее расщепление: 14 синих, 10 зеленых и 2 желтых. Что сначала приходит в голову? 7:5:1. Но это неправильный ход рассуждений. Почему? Потому что для подсчета соотношений в F2 существуют определенные правила. Для начала необходимо предположить количество генов, участвующих в определении признака. Как это сделать? Для этого нужно посмотреть, сколько фенотипических классов образовалось в F2 и каково их примерное соотношение. Если образовалось более 3-х классов, то можно с уверенностью говорить, что в определении признака участвует 2 и более генов. Однако наличие 2-3 классов в F2 также не всегда говорит о том, что это разные аллельные варианты одного гена (конечно, если это не 3:1 или 1:2:1). Как правильно посчитать соотношение? Сначала нужно определить (предположить) примерное количество генов, участвующих в определении признака. Далее нужно подсчитать количество возможных вариантов генотипов, которые образуются в F2. Это делается по формуле: Х= 4 в степени "n" (таблица 1), где n - предполагаемое количество генов, а Х - количество возможных вариантов генотипов. В нашем примере мы предполагаем, что в наследовании признака окраса участвуют 2 гена, значит Х=16. Действительно, по количеству мальков в трех фенотипических классах (синие, красные и желтые) расщепление отличается от неполного доминирования (1:2:1). Общее кол-во рыб в F2 (а также отдельное количество самок и самцов) делится на 16 (14+10+2 / 16) и мы получаем 1/16 часть = 1,6. Учитывая, что возможных вариантов генотипов может быть 16, мы должны разделить количество рыб в каждом классе на 1,6: синие - 14/1,6=8,7; зеленые - 10/1,6=6,2; желтые - 2/1,6=1,2, что примерно равно отношению 9:6:1. Мы получили истинное расщепление 9:6:1, а не 7:5:1. Возможное сочетания гамет и генотипов поможет выявить так называемая решетка Пеннета, используя которую можно легко понять взаимодействие аллелей двух генов. По вертикали в ней расписаны гаметы самца, а по горизонтали – самки. Таким образом, в клетках решетки показаны все возможные варианты генотипов (рис. 4).
 Рис. 4. Решетка Пеннета (для двух генов - А и B).
Одно НО. Мы расписали решение этой задачи на маленькой выборке размером всего 26 рыб, при условии менделеевского расщепления и отсутствия взаимодействия генов между собой. А что было бы, если мы анализировали выборку размером в 5 или 10 раз больше – 130 и 260 рыб, соответственно. При этом генов было бы больше, чем 2 и они взаимодействовали между собой. В этом случае результат был бы более корректный. Поэтому важно отметить, что серьезный генетический анализ правильнее проводить на большой выборке. Только так вы сможете достаточно точно определить генетику рыб. Например, во втором поколении, могло получиться не 4, а гораздо большее (до 16!!!) количество классов за счет как минимум 4 пигментных генов, а также их взаимодействия. Поэтому приведенный выше пример корректен для анализа различия именно по 2 генам.
В заключении статьи рассмотрим несколько ключевых взаимодействий между двумя генами с разной локализацией (расположением в хромосомах). При локализации гена в половых хромосомах результаты скрещиваний будут отличаться от классического менделевского наследования.
При локализации признака в Х-хромосомев первом поколении одного из реципрокных скрещиваний (когда одновременно проводится два скрещивания, но в одном случае носителем изучаемого аллеля (признака) является самка, а в другом – самец) проявится крисс-кросс наследование (признак от матери перейдет к сыну, а от отца - к дочери). В другом реципрокном скрещивании наблюдается единообразие - у самок и самцов признак проявляется одинаково. При этом во втором поколении расщепление будет 3:1 и 1:1, соответственно.
Если ген локализован в Y-хромосоме, то он передается только самцам, независимо от того, доминантный признак или рецессивный.
В случае частичного сцепления с полом, когда ген находится в обеих половых хромосомах (XY-сцепленность), в F1 наблюдается единообразие и только во втором поколении (F2) частичное сцепление с полом обнаруживается только по результатам реципрокных скрещиваний. В обоих скрещиваниях расщепления будут одинаковыми (3:1), однако в одном их них рецессивный признак (аллель) проявится только у самок, а в другом – у самцов. Для этого достаточно расписать гаметы родителей в решетке Пеннета.
Более сложным для генетического анализа является наследование признаков, когда один из генов находится в аутосоме, а другой – в Х-хромосоме (взаимодействие генов). В первом поколении реципрокных скрещиваний в одном случае будет единообразие (если в Х-хромосоме и аутосоме самки доминантные аллели), в обратном скрещивании (когда в X-хромосоме и аутосоме самки находятся рецессивные аллели) наблюдается расщепление – самки будут нести доминантные аллели изучаемых генов, а самцы будут гетерозиготны по аутосомальному гену и нести рецессивный аллель в Х-хромосоме. В F2 при этом в первом случае по аутосомальному гену у самцов и самок будет наблюдаться расщепление 3:1, а по гену Х-хромосомы также 3:1, но особи с рецессивным аллелем будут только мужского пола (у самок не будет происходить расщепление). В обратном скрещивании по обоим изучаемым генам у самцов и самок будут расщепления 3:1 и 1:1, соответственно.
Безусловно, рассказать в одной статье обо всех премудростях генетики и селекции рыб просто невозможно. Можно лишь обозначить основные направления, разобрать азы этой науки и совсем немного приоткрыть занавес непонимания аквариумистов в этом вопросе. Надеюсь, что мне это удалось.
Желаю всем аквариумистам успехов в генетике и селекции аквариумных рыб!!!
Литература
1. И.В. Болгова. «Сборник задач по общей биологии с решениями». М. - Изд. «Оникс». -2008.
2. Н.Н. Орлова. «Генетический анализ». М.- Изд. «МГУ». -1991.
3. С.Г. Инге-Вечтомов. «Генетика с основами селекции». М.- Изд. «Высшая школа». – 1989.
4. Журнал «Генетика гуппи». Под ред. В.В. Сторожева, С.А. Апрятина.- №№ 1, 2.- 2009; №1.- 2010.
5. У.Клаг, М.Каммингс. «Основы генетики». М.- Изд. «Техносфера». – 2007.
© С.А. Апрятин, 2010
© "Аквариум", 2010
Рис. 4. Решетка Пеннета (для двух генов - А и B).
Одно НО. Мы расписали решение этой задачи на маленькой выборке размером всего 26 рыб, при условии менделеевского расщепления и отсутствия взаимодействия генов между собой. А что было бы, если мы анализировали выборку размером в 5 или 10 раз больше – 130 и 260 рыб, соответственно. При этом генов было бы больше, чем 2 и они взаимодействовали между собой. В этом случае результат был бы более корректный. Поэтому важно отметить, что серьезный генетический анализ правильнее проводить на большой выборке. Только так вы сможете достаточно точно определить генетику рыб. Например, во втором поколении, могло получиться не 4, а гораздо большее (до 16!!!) количество классов за счет как минимум 4 пигментных генов, а также их взаимодействия. Поэтому приведенный выше пример корректен для анализа различия именно по 2 генам.
В заключении статьи рассмотрим несколько ключевых взаимодействий между двумя генами с разной локализацией (расположением в хромосомах). При локализации гена в половых хромосомах результаты скрещиваний будут отличаться от классического менделевского наследования.
При локализации признака в Х-хромосомев первом поколении одного из реципрокных скрещиваний (когда одновременно проводится два скрещивания, но в одном случае носителем изучаемого аллеля (признака) является самка, а в другом – самец) проявится крисс-кросс наследование (признак от матери перейдет к сыну, а от отца - к дочери). В другом реципрокном скрещивании наблюдается единообразие - у самок и самцов признак проявляется одинаково. При этом во втором поколении расщепление будет 3:1 и 1:1, соответственно.
Если ген локализован в Y-хромосоме, то он передается только самцам, независимо от того, доминантный признак или рецессивный.
В случае частичного сцепления с полом, когда ген находится в обеих половых хромосомах (XY-сцепленность), в F1 наблюдается единообразие и только во втором поколении (F2) частичное сцепление с полом обнаруживается только по результатам реципрокных скрещиваний. В обоих скрещиваниях расщепления будут одинаковыми (3:1), однако в одном их них рецессивный признак (аллель) проявится только у самок, а в другом – у самцов. Для этого достаточно расписать гаметы родителей в решетке Пеннета.
Более сложным для генетического анализа является наследование признаков, когда один из генов находится в аутосоме, а другой – в Х-хромосоме (взаимодействие генов). В первом поколении реципрокных скрещиваний в одном случае будет единообразие (если в Х-хромосоме и аутосоме самки доминантные аллели), в обратном скрещивании (когда в X-хромосоме и аутосоме самки находятся рецессивные аллели) наблюдается расщепление – самки будут нести доминантные аллели изучаемых генов, а самцы будут гетерозиготны по аутосомальному гену и нести рецессивный аллель в Х-хромосоме. В F2 при этом в первом случае по аутосомальному гену у самцов и самок будет наблюдаться расщепление 3:1, а по гену Х-хромосомы также 3:1, но особи с рецессивным аллелем будут только мужского пола (у самок не будет происходить расщепление). В обратном скрещивании по обоим изучаемым генам у самцов и самок будут расщепления 3:1 и 1:1, соответственно.
Безусловно, рассказать в одной статье обо всех премудростях генетики и селекции рыб просто невозможно. Можно лишь обозначить основные направления, разобрать азы этой науки и совсем немного приоткрыть занавес непонимания аквариумистов в этом вопросе. Надеюсь, что мне это удалось.
Желаю всем аквариумистам успехов в генетике и селекции аквариумных рыб!!!
Литература
1. И.В. Болгова. «Сборник задач по общей биологии с решениями». М. - Изд. «Оникс». -2008.
2. Н.Н. Орлова. «Генетический анализ». М.- Изд. «МГУ». -1991.
3. С.Г. Инге-Вечтомов. «Генетика с основами селекции». М.- Изд. «Высшая школа». – 1989.
4. Журнал «Генетика гуппи». Под ред. В.В. Сторожева, С.А. Апрятина.- №№ 1, 2.- 2009; №1.- 2010.
5. У.Клаг, М.Каммингс. «Основы генетики». М.- Изд. «Техносфера». – 2007.
© С.А. Апрятин, 2010
© "Аквариум", 2010
|
|
Поиск по сайту:
|