Слуховая система: кодирование частоты звука
Слуховая система: кодирование частоты звука
Соматосенсорная система анализирует стимулы на основе их расположения на карте тела. Благодаря аналогичному процессу, зрительные стимулы являются ретинотопически картированными и анализируются на предмет их местоположения в визуальном (пространственном) мире. Однако слуховая система использует совершенно иной метод анализа. Слуховая периферия, или улитка, картирует звуковые стимулы скорее по их частотному содержанию, чем по пространственному расположению, и последующие стадии слуховой системы используют этот тонотопический анализ, чтобы определить смысл речи и значение других сложных звуков. Как частота звука кодируется волосковыми клетками улитки, и как эти сигналы обрабатываются на более высоких уровнях слуховой системы? Как определяется пространственное расположение звука?
У наземных позвоночных звуковые волны в воздухе входят во внешнее ухо, колеблют барабанную перепонку и, благодаря серии механических сопряжений в среднем ухе, конвертируются в колебания жидкости улитки. Колебания жидкости, в свою очередь, вызывают вибрацию базилярной мембраны, на которой находятся сенсорные волосковые клетки в составе органа Корти. Этот процесс описан поэтически Олдосом Хаксли:
Трубный звук Понгилеони и скрежет безымянных скрипок потрясли воздух в большой зале, вызвали дрожь в оконных стеклах, и это, в свою очередь, сотрясло воздух в апартаментах лорда Эдварда на дальней стороне. Сотрясающийся воздух задребезжал в барабанных перепонках лорда Эдварда; соединенные между собой косточки молоточка (ушной косточки), наковальни внутреннего уха и стремени (слуховой кости среднего уха) были приведены в движение, так что возбудили мембрану окна (oval window) и подняли бесконечно малую бурю в жидкости лабиринта. Волосковые окончания слухового нерва дрожали, как водоросли в бушующем море; огромное количество непонятных чудес произошло в мозгу, и лорд Эдвард в экстазе прошептал: «Бах!»
Улитка
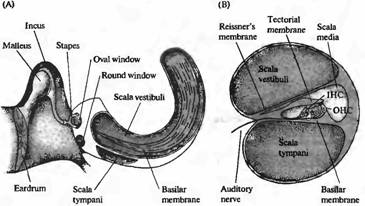
Канал улитки разделен на три отдела. Средняя лестница (scala media) содержит раствор с высоким содержанием калия, эндолимфу. Он отделен от покрывающей его вестибулярной лестницы (scala vestibuli) Рейсснеровой мембраной, а от наполненного жидкостью пространства барабанной лестницы (scala timpani) — тесными межклеточными соединениями между апикальными окончаниями волосковых клеток и окружающими вспомогательными клетками (рис. 1В). Барабанная и вестибулярная лестницы содержат перилимфу, сходную по составу с цереброспинальной жидкостью; средняя лестница содержит эндолимфу, ионный состав которой похож на состав цитоплазмы: высокое содержание калия, низкое — натрия и с концентрацией кальция, поддерживаемой на микромолярном уровне. Необычный состав внеклеточной жидкости устанавливается благодаря работе ионных насосов в клетках, расположенных в сосудистом слое (stria vascularis), т. е. в секреторном эпителии, выстилающем боковую стенку средней лестницы.
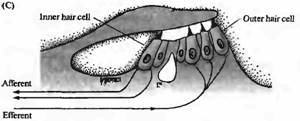
Существуют две различные группы волосковых клеток в улитке млекопитающих: внутренние волосковые клетки и внешние волосковые клетки. Они различаются по своему расположению (внутренние волосковые клетки ближе к центральной оси спирали улитки) и по паттерну иннервации. Внутренние волосковые клетки получают более 90% афферентных контактов улитки; внешние волосковые клетки являются постсинаптическими мишенями для эфферентной иннервации (рис. 1С). Эти различия в паттернах иннервации ставят интересные вопросы, касающиеся функциональной роли этих двух типов клеток, которые будут обсуждаться далее в этой главе. Покровная мембрана (tectorial membrane) налегает на волосковые пучки как внутренних, так и внешних волосковых клеток, и различие в движении покровной и базилярной мембран приводит к боковому сдвигу, открывающему каналы механотрансдукции.
Частотная избирательность: механическая настройка
Звуковое восприятие — это не просто вопрос выявления звукового стимула. Информационное содержание звука является функцией его частотного состава. Таким образом, слуховая система зависит от способности ее механосенсорных волосковых клеток быть частотно избирательными. В улитке млекопитающих настройка по частоте зависит от механических свойств специализированной структуры, в которой находятся сенсорные клетки, аналогично механической адаптации, осуществляемой многослойной капсулой в тельце Пачини. Ширина и толщина базилярной мембраны закономерно меняется по длине канала улитки (рис. 1 А). У основания улитки (ближе к овальному окну) базилярная мембрана узкая и жесткая; у противоположного конца (вершина улитки) она широкая и гибкая. По причине этой изменяющейся жесткости входящие звуковые волны вызывают движение жидкости и вибрацию базилярной мембраны в разных участках в зависимости от частоты звука.
Это свойство функции улитки было описано фон Бекеси, который использовал стробоскопическое освещение отражающих частиц, рассеянных в улитке, чтобы визуализировать паттерн вибрации. Он обнаружил, что звуки высокой частоты вызывают максимальные колебания на более толстой, жест кой нижней части мембраны, а звуки низкой частоты вызывают максимальную вибрацию на более гибкой верхней части мембраны улитки. Фон Бекеси назвал этот паттерн колебаний «бегущей волной».
Как следствие механической настройки базилярной мембраны,
|
| Рис. 1. Структура улитки. (А) Схема среднего уха и улитки, показывающая систему косточек, с помощью которых барабанная перепонка соединена с мембраной овального окна улитки. Улитка дана рассеченной и не скрученной, чтобы показать ее основные внутренние полости (вестибулярную и барабанную лестницы) и форму базилярной мембраны, расширяющейся от основания улитки к ее вершине. (В) Поперечный разрез улитки, показывающий структурные взаимоотношения между базилярной мембраной, внутренними (IHC) и наружными (ОНС) волосковыми клетками и покровной мембраной. (С) Иннервация волосковых клеток улитки. Около 95 % афферентных волокон являются постсинаптическими мишенями для IHC. До 20 афферентных волокон контактирует с одиночной IHC. ОНС имеют мало афферентных связей, зато являются постсинаптическими мишенями для холинергических эфферентных нейронов, которые проецируются из оливарного комплекса, расположенного в стволе мозга. |
волосковые клетки и их постсинаптические афферентные волокна у основания улитки преимущественно стимулируются высокочастотным звуком, тогда как те, что находятся у вершины улитки, лучше всего реагируют на низкочастотный звук (рис. 2). Частотная избирательность афферентов улитки может быть измерена при помощи отведения потенциалов действия, возникающих в ответ на чистые тона звука,
| Рис. 2. Настройка улитки. (А) Локализация участков улитки, в которых звуковые волны вызывают максимальное смещение базилярной мембраны, зависит от частоты. Кривые представляют относительное смещение на указанных частотах (100-2000 Гц). (В) Типичные кривые настройки четырех индивидуальных волокон слухового нерва, иннервирующих различные участки улитки. Показана зависимость между частотой звукового стимула и интенсивностью звука в децибелах (dB) необходимой для возникновения разряда в волокне. Пики соответствуют оптимальным звуковым частотам (т. е. частотам, при которых интенсивность стимула для каждого волокна минимальна). against frequency of the auditory stimulus. | 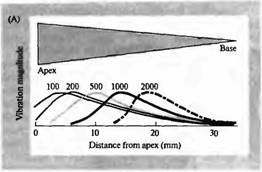

|
частота и интенсивность которых систематически варьирует. Результирующая кривая настройки V-образна, с оптимальной, или характеристической, частотой, определяемой как чистый тон, к которому волокно наиболее чувствительно. Характеристическая частота каждого волокна определяется по тому, где именно в канале улитки оно контактирует с волосковой клеткой.
Звуковая частота картируется на протяжении эпителиального массива рецепторов, производящих кохлеотопическую, или тонотопическую карту частоты. Поэтому, слуховая система может использовать принципы нейронной обработки — такие, как латеральное торможение — основанные на тонотопии, чтобы истолковывать и обрабатывать информацию, закодированную в звуковой частоте таким же образом, как соматотопические карты лежат в основе обработки соматосенсорных сигналов.