Зрительный бугор (таламус) и подбугорье (гипоталамус) являются частями промежуточного мозга. Импульсы от всех рецепторов организма поступают в ядра таламуса. Поступившая информация в таламусе перерабатывается и направляется к большим полушариям мозга. Таламус соединяется с мозжечком и т.н. лимбической системой. Гипоталамус регулирует вегетативные функции организма. Влияние гипоталамуса осуществляется через нервную систему и железы внутренней секреции. Гипоталамус также участвует в регуляции функций многих эндокринных желез и обмена веществ, а также в регуляции температуры тела и деятельности сердечно-сосудистой и пищеварительной систем.
Лимбическая система
В формировании эмоционального поведения человека большую роль играет лимбическая система. К лимбической системе относят нервные образования головного мозга, расположенные на срединной стороне конечного мозга, около верхнего ствола мозга. Эта часть головного мозга еще не вполне изучена. Предполагается, что лимбическая система и управляемое ею подбугорье являются ответственными за множество наших чувств и желаний, например, под их воздействием возникают жажда и голод, страх, агрессивность, половое влечение.
Функции ствола головного мозга
Ствол головного мозга - это филогенетически древняя часть мозга, состоящая из среднего, заднего и продолговатого мозга. В среднем мозге имеются первичные зрительные и слуховые центры. С их участием осуществляются ориентировочные рефлексы на свет и звук. В продолговатом мозге расположены центры регуляции дыхания, сердечно-сосудистой деятельности, функций пищеварительных органов, а также обмена веществ. Продолговатый мозг принимает участие в осуществлении таких рефлекторных актов, как жевание, сосание, чихание, глотание, рвота.
|
|
Функции мозжечка
Мозжечок контролирует движения тела. К мозжечку приходят импульсы от всех рецепторов, которые раздражаются во время движений тела. Функция мозжечка может нарушаться при принятии алкоголя или других веществ, вызывающих головокружение. Поэтому под действием опьянения люди не способны нормально координировать свои движения. В последние годы появляется все больше доказательств, что мозжечок имеет значение и в познавательной деятельности человека.
Черепно-мозговые нервы
Помимо спинного мозга очень важны и двенадцать черепно-мозговых нервов: I и II пары -обонятельный и зрительный нервы; III, IV VI пары - глазодвигательные нервы; V пара -тройничный нерв - иннервирует жевательные мышцы; VII - лицевой нерв - иннервирует мимические мышцы, содержит также секреторные волокна к слезной и слюнным железам; VIII пара - преддверно-улитковый нерв - связывает органы слуха, равновесия и гравитации; IX пара - языкоглоточный нерв — иннервирует глотку, ее мышцы, околоушную железу, вкусовые почки языка; X пара - блуждающий нерв -разделяется на ряд ветвей, которые иннервируют легкие, сердце, кишечник, регулируют их функции; XI пара - добавочный нерв - иннервирует мышцы плечевого пояса. В результате слияния спинномозговых нервов образуется XII пара - подъязычный нерв - иннервирует мышцы языка и подъязычный аппарат.
Развитие и дифференциация структур Нервная система у высокоорганизованных животных обусловили её разделение на соматическую и вегетативную нервную систему. Особенность строения вегетативной Нервная система та, что её волокна, отходящие от ЦНС, не доходят непосредственно до эффектора, а сначала вступают в периферические ганглии, где оканчиваются на клетках, отдающих аксоны уже непосредственно на иннервируемый орган. В зависимости от того, где расположены ганглии вегетативной Нервная система, и некоторых её функциональных особенностей вегетативную Нервная система делят на 2 части: парасимпатическую и симпатическую.
|
|
Структурная и функциональная единица Нервная система - нейрон, состоящий из тела нервной клетки и отростков - аксона и дендритов. Кроме нервных клеток, в структуру Нервная система входят глиальные клетки. Нейроны являются в известной мере самостоятельными единицами - их протоплазма не переходит из одного нейрона в другой. Взаимодействие между нейронами осуществляется благодаря контактам между ними - синапсам. В области контакта между окончанием одного нейрона и поверхностью другого в большинстве случаев сохраняется особое пространство - синаптическая щель - шириной в несколько сот ангстрем. Основные функции нейронов: восприятие раздражений, их переработка, передача этой информации и формирование ответной реакции. В зависимости от типа и хода нервных отростков (волокон), а также их функций нейроны подразделяют на: а) рецепторные (афферентные), волокна которых проводят нервные импульсы от рецепторов в ЦНС; тела их находятся в спинальных ганглиях или ганглиях черепномозговых нервов; б) двигательные (эфферентные), связывающие ЦНС с эффекторами; тела и дендриты их находятся в ЦНС, а аксоны выходят за её пределы (за исключением эфферентных нейронов вегетативной Нервная система, тела которых расположены в периферических ганглиях); в) вставочные (ассоциативные) нейроны, служащие связующими звеньями между афферентными и эфферентными нейронами; тела и отростки их расположены в ЦНС.
|
|
Деятельность Нервная система основывается на двух процессах: возбуждении и торможении. Возбуждение может быть распространяющимся (см. Импульс нервный)или местным - нераспространяющимся, стационарным (последнее открыто Н. Е. Введенским в 1901) Торможение - процесс, тесно связанный с возбуждением и внешне выражающийся в снижении возбудимости клеток. Одна из характерных черт тормозного процесса - отсутствие способности к активному распространению по нервным структурам (явление торможения в нервных центрах впервые было установлено И. М. Сеченовым в 1863).
Клеточные механизмы возбуждения и торможения подробно изучены. Тело и отростки нервной клетки покрыты мембраной, постоянно несущей на себе разность потенциалов (так называемый мембранный потенциал). Раздражение расположенных на периферии чувствительных окончаний афферентного нейрона преобразуется в изменение этой разности потенциалов (см. Биоэлектрические потенциалы). Возникающий вследствие этого нервный импульс распространяется по нервному волокну и достигает его пресинаптического окончания, где вызывает выделение в синаптическую щель высокоактивного химического вещества - медиатора. Под влиянием последнего в постсинаптической мембране, чувствительной к действию медиатора, происходит молекулярная реорганизация поверхности. В результате постсинаптическая мембрана начинает пропускать ионы и деполяризуется, вследствие чего на ней возникает электрическая реакция в виде местного возбуждающего постсинаптического потенциала (ВПСП), вновь генерирующего распространяющийся импульс. Нервные импульсы, возникающие при возбуждении особых тормозящих нейронов, вызывают гиперполяризацию постсинаптической мембраны и, соответственно, тормозящий постсинаптический потенциал (ТПСП). Помимо этого, установлен и др. вид торможения, формирующийся в пресинаптической структуре, - пресинаптическое торможение, обусловливающее длительное снижение эффективности синаптической передачи. В основе деятельности Нервная система лежит рефлекс, т. е. реакция организма на раздражения рецепторов, осуществляемая при посредстве Нервная система Термин «рефлекс» был впервые введён в зарождавшуюся физиологию Р. Декартом в 1649, хотя конкретных представлений о том, как осуществляется рефлекторная деятельность, в то время ещё не было. Такие сведения были получены лишь значительно позже, когда морфологи приступили к исследованию строения и функций нервных клеток. Все рефлекторные процессы связаны с распространением возбуждения по определённым нервным структурам - рефлекторным дугам. Основные элементы рефлекторной дуги: рецепторы, центростремительный (афферентный) нервный путь, внутрицентральные структуры различной сложности, центробежный (эфферентный) нервный путь и исполнительный орган (эффектор). Различные группы рецепторов возбуждаются раздражителями разной модальности (т. е. качественной специфичности) и воспринимают раздражения, исходящие как из внешней среды (экстерорецепторы - органы зрения, слуха, обоняния и др.), так и из внутренней среды организма (интерорецепторы, возбуждающиеся при механических, химических, температурных и др. раздражениях внутренних органов, мышц и др.). Нервные сигналы, несущие в ЦНС информацию от рецепторов по нервным волокнам, лишены модальности и обычно передаются в виде серии однородных импульсов. Информация о различных характеристиках раздражений кодируется изменениями частоты импульсов, а также приуроченностью нервной импульсации к определённым волокнам (так называемое пространственно-временное кодирование). Совокупность рецепторов данной области тела животного или человека, раздражение которых вызывает определённый тип рефлекторной реакции, называется рецептивным полем рефлекса. Такие поля могут накладываться друг на друга. Совокупность нервных образований, сосредоточенных в ЦНС и ответственных за осуществление данного рефлекторного акта, обозначают термином нервный центр. На отдельном нейроне в Нервная система может сходиться огромное число окончаний волокон, несущих импульсы от др. нервных клеток. В каждый данный момент в результате сложной синаптической переработки этого потока импульсов обеспечивается дальнейшее проведение лишь одного, определённого сигнала - принцип конвергенции, лежащий в основе деятельности всех уровней Нервная система («принцип конечного общего пути» Шеррингтона, получивший развитие в трудах Ухтомского и др.). Пространственно-временная суммация синаптических процессов служит основой для различных форм избирательного функционального объединения нервных клеток, лежащего в основе анализа поступающей в Нервная система информации и выработки затем команд для выполнения различных ответных реакций организма. Такие команды, как и афферентные сигналы, передаются от одной клетки к другой и от ЦНС к исполнительным органам в виде последовательностей нервных импульсов, возникающих в клетке в том случае, когда суммирующиеся возбуждающие и тормозящие синаптические процессы достигают определённого (критического для данной клетки) уровня - порога возбуждения. Несмотря на наследственно закрепленный характер связей в основных рефлекторных дугах, характер рефлекторной реакции может в значительной степени изменяться в зависимости от состояния центральных образований, через которые они осуществляются. Так, резкое повышение или понижение возбудимости центральных структур рефлекторной дуги может не только количественно изменить реакцию, но и привести к определённым качественным изменениям в характере рефлекса. Примером такого изменения может служить явление доминанты. Важное значение для нормального протекания рефлекторной деятельности имеет механизм так называемой обратной афферентации - информации о результате выполнения данной рефлекторной реакции, поступающей по афферентным путям от исполнительных органов. На основании этих сведений в случае, если результат неудовлетворителен, в сформировавшейся функциональной системе могут происходить перестройки деятельности отдельных элементов до тех пор, пока результат не станет соответствовать уровню, необходимому для организма (П. К. Анохин, 1935). Всю совокупность рефлекторных реакций организма делят на две основные группы: безусловные рефлексы - врождённые, осуществляемые по наследственно закрепленным нервным путям, и условные рефлексы, приобретённые в течение индивидуальной жизни организма путём образования в ЦНС временных связей. Способность образования таких связей присуща лишь высшему для данного вида животных отделу Нервная система (для млекопитающих и человека - это кора головного мозга). Образование условнорефлекторных связей позволяет организму наиболее совершенно и тонко приспосабливаться к постоянно изменяющимся условиям существования. Условные рефлексы были открыты и изучены И. П. Павловым в конце 19 - начале 20 вв. Исследование условнорефлекторной деятельности животных и человека привело его к созданию учения о высшей нервной деятельности (ВНД) и анализаторах. Каждый анализатор состоит из воспринимающей части - рецептора, проводящих путей и анализирующих структур ЦНС, обязательно включающих её высший отдел. Кора головного мозга у высших животных - совокупность корковых концов анализаторов; она осуществляет высшие формы анализаторной и интегративной деятельности, обеспечивая совершеннейшие и тончайшие формы взаимодействия организма с внешней средой. Нервная система обладает способностью не только немедленно перерабатывать поступающую в неё информацию при помощи механизма взаимодействующих синаптических процессов, но и хранить следы прошлой активности (механизмы памяти). Клеточные механизмы сохранения в высших отделах Нервная система длительных следов нервных процессов, лежащие в основе памяти, интенсивно изучаются. Наряду с перечисленными выше функциями Нервная система осуществляет также регулирующие влияния на обменные процессы в тканях - адаптационно-трофическую функцию (И. П. Павлов, Л. А. Орбели, А. В. Тонких и др.). При перерезке или повреждении нервных волокон свойства иннервируемых ими клеток изменяются (это касается как физико-химических свойств поверхностной мембраны, так и биохимических процессов в протоплазме), что, в свою очередь, сопровождается глубокими нарушениями в состоянии органов и тканей (например, трофическими язвами). Если иннервация восстанавливается (в связи с регенерацией нервных волокон), то указанные нарушения могут исчезнуть.
Эволюция кровеносной системы У многих беспозвоночных животных сосудистой системы как таковой нет. В частности, у губок, кишечнополостных и плоских червей транспорт питательных веществ и кислорода в разные части тела осуществляется путем диффузии их тканевых жидкостей. Однако у организмов других групп появляются сосуды, обеспечивающие циркуляцию тканевой жидкости в определенных направлениях. В начале развиваются примитивные сосуды, но в результате развития в сосудах мышечной ткани они в дальнейшем оказываются способными к сокращению. Параллельно развивается в кровь жидкость, заполняющая сосуды.
Кровеносная система впервые встречается (развивается) у кольчатых червей, у которых она является замкнутой, представляя собой систему, состоящую из верхнего спинного и нижнего брюшного сосудов, объединенных кольцевыми сосудами, опоясывающими кишечник. Кроме того, от спинного и брюшного сосудов отходят более мелкие сосуды к стенкам тела. К головному отделу кровь идет по спинной стороне, а к заднему -- по брюшной.
У членистоногих, кровеносная система не замкнута, т. к. сосуды открываются в полость тела. Спинной сосуд разделяется перегородками (клапанами) на отдельные камеры -- сердца, сокращения которых заставляют проходить кровь в артерии, а из последних -- в пространства между органами. Из этих пространств кровь затем поступает в околосердечную полость.
У моллюсков кровеносная система тоже не замкнута и представлена сердцем, состоящим из нескольких предсердий и желудочка, а также артериальными и венозными сосудами. Вены впадают в предсердие, тогда как артерии отходят от желудочка.
Кровь у беспозвоночных несет кислород к тканям, удаляет двуокись углерода и транспортирует питательные вещества, гормоны, а также шлаковые продукты азотистого обмена. Кровь беспозвоночных по сравнению с кровью позвоночных содержит меньшее количество белков и форменных элементов. Единственным форменным элементом крови большинства беспозвоночных являются лейкоциты. Поэтому очень простой состав крови у беспозвоночных компенсируется разными способами транспорта кислорода, при которых используются гемоглобин, гемоцианин или другие дыхательные пигменты.
Предполагают, что развитие гемоглобина беспозвоночных шло независимо от развития гемоглобина позвоночных, поскольку этот пигмент у них обладает несколько отличной функцией. В частности, у беспозвоночных он выполняет запасательную функцию, обеспечивая защиту против недостатка кислорода в неблагоприятной среде.
По мере усложнения организации живых существ отмечается повышение количества кислорода в крови, т. е. повышается кислородная способность дыхательных пигментов.
Считают, что гемоглобин имеет древний возраст. Поскольку некоторые бактерии синтезируют гемопротеин, содержащий участки с аминокислотным составом, близким к последовательностям глобинов высших эукариотов, то предполагают, что гемоглобин впервые возник еще у прокариот. Однако не исключено, что у эукариот и прокариот имела место конвергентная эволюция.
Гемоглобин встречается у плоских, круглых и кольчатых червей, членистоногих, моллюсков, иглокожих, рыб, земноводных, рептилий, птиц, млекопитающих. Молекулярная масса гемоглобина беспозвоночных колеблется в пределах 17 000-2 750 000. У многих членистоногих и моллюсков встречается гемопианин, представляющий собой медьсодержащий белок молекулярной массой 400 000--5 000 000. Дыхательный пигмент обеспечивает у беспозвоночных буферную способность крови. Кроме того он ответственен и за осмотический баланс кровяной жидкости. Большинство насекомых не содержит совсем дыхательного пигмента, имея в крови растворенный кислород.
У полухордовых кровеносные сосуды еще не выстланы эндотелием. Сердце урохордовых представляет собой мускульное выпячивание около желудка. У хордовых кровеносная система характеризуется дальнейшим совершенствованием. У них развиваются сердце, сосуды, кровь.
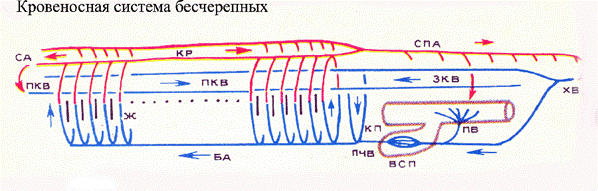
У ланцетника кровеносная система является замкнутой, но сердца еще нет; его функцию выполняет передний отдел крупного сосуда в виде брюшной аорты. Впервые сердце появляется у водных позвоночных. В частности, у рыб сердце двухкамерное с предсердием и желудочком.
Круг кровообращения один, в котором артериальная и венозная кровь не смешиваются. Из сердца венозная кровь идет к жабрам, где, окисляясь, она становится артериальной, после чего расходится по артериям ко всем частям тела. К сердцу кровь вновь доставляется венами. У пресмыкающихся желудочек разделен неполной перегородкой на правую (венозную) и левую (артериальную) половины. У крокодилов желудочек разделен полностью. Однако у пресмыкающихся разделения артериального и венозного тока крови еще не происходит.
У наземных позвоночных развивается трех-, а затем четырехкамерное сердце, что явилось результатом очень крупного ароморфоза. У птиц и млекопитающих сердце построено из мышц и разделено на четыре камеры в виде двух предсердий и двух желудочков Имеются два круга кровообращения, один из которых большой, второй малый. Благодаря этим кругам артериальная и венозная кровь не смешиваются. Артериальная кровь выходит из сердца, венозная вступает в сердце.
Сердце позвоночных относят к миогенному типу, поскольку его сокращения начинаются внутри за счет сократительных элементов, называемых миофибриллами. У рыб, земноводных и пресмыкающихся сокращения сердца начинаются в тонкой мышечной стенке венозного синуса и продолжаются посредством внутренней проводящей системы через предсердие, потом к желудочкам. У птиц и млекопитающих венозного синуса нет.
У позвоночных усложняются строение и функции крови. Кровь состоит из форменных элементов и плазмы, увеличивается их масса. Например, в крови млекопитающих клеточная масса крови составляет 45%, плазменная -- 55%. Гемоглобин содержится в эритроцитах, которые у всех позвоночных, кроме млекопитающих, содержат ядра. У земноводных клетки крови очень крупные по сравнению с пресмыкающимися, птицами и млекопитающими. В 1 мл крови человека содержится 5 млн эритроцитов и 8--10 тыс лейкоцитов. Группы крови современных людей также являются результатом их эволюции В плазме крови содержатся ферменты, гормоны и другие соединения, необходимые для метаболизма клеток.
В ходе эволюции у высших животных взаимосвязи обмена веществ разных органов, обеспечиваемые кровью, получили дальнейшее усложнение. Одна из важнейших функций крови заключается в транспорте кислорода от легких к тканям и в транспорте СО2, являющимся конечным продуктом дыхания, от тканей к легким, причем эта функция крови развивалась в направлении способности транспортировать большие объемы кислорода. Основная часть кислорода, потребляемого организмом человека, транспортируется гемоглобином эритроцитов, масса которого составляет примерно треть массы эритроцитов. Гемоглобин представляет собой очень сложный белок, состоящий из двух а-полипептидных цепей и двух -полипептидных цепей, каждый из которых присоединяет по одной гемовой группе. Его происхождение является очень древним
Эритроциты участвуют также в транспорте СО2 из тканей в легкие, откуда он выводится в процессе выдыхания. Эволюционным приобретением млекопитающих, в том числе человека, является то, что их гемоглобин приспособлен как для транспорта кислорода, так и для транспорта СОд, причем оба эти транспорта взаимоусиливаемы.
Кровь ответственна и за перенос питательных веществ из тонкого кишечника в печень и другие органы, а также за выведение шлаков из тканей в почки. Эти функции тоже развивались в направлении увеличения количественной способности в связи с увеличивающейся интенсификацией метаболизма по мере усложнения организации живых существ.
Диффузия кислорода, двуокиси углерода, питательных веществ и метаболитов происходит лишь в капиллярах.
Параллельно с развитием кровеносной системы шло развитие лимфатической системы, которая впервые появилась у хордовых. Эта система в ходе эволюции обособилась из венозной системы.
Лимфатическая система состоит из лимфатических сосудов, лимфатических узлов и лимфы.
Лимфатические сосуды построены из эластичной и мышечной тканей и выстланы внутри эндотелием. Для них характерен такой же внутренний объем, как и у кровеносных капилляров. Лимфатические сосуды очень разветвлены. Они в виде очень тонких капилляров начинаются с межклеточных пространств, формируя затем более крупные сосуды. С помощью лимфатических сосудов лимфа проходит от тканей в венозное русло. Все лимфатические сосуды сливаются в один общий сборный сосуд (млечный путь). Одиночные лимфатические узлы обнаруживаются в толще слизистой оболочки и подслизистой основы кишечника у рыб, земноводных, пресмыкающихся, птиц и млекопитающих, но групповые лимфатические узлы развиваются лишь у млекопитающих.
У позвоночных по мере усложнения их организации наступает приближение лимфатических капилляров к клеткам и лимфоид-ным узелкам. Например, расстояние между лимфатическими капиллярами, кишечными эпителиоцитами и лимфоидными узелками у рыб составляет 150-180 мкм, но затем прогрессивно уменьшается у земноводных, пресмыкающихся и птиц, составляя у млекопитающих всего лишь 50-30 мкм.
В ходе эволюции многоклеточных животных кровеносная система сформировалась на месте рудиментов первичной полости тела, вытесненной у высших животных вторичной полостью тела, или целомом. Кровеносная система приняла на себя в организме высших животных универсальную транспортную роль передвижение питательных веществ, усвоенных органами пищеварения, и кислорода от органов дыхания ко всем органам тела, перенос продуктов метаболизма к органам выделения, углекислоты — к органам дыхания, доставка гормонов от эндокринных желез к органам, где данные гормоны будут действовать. В силу этого кровеносная система стала и одной из важнейших интегрирующих систем организма, обеспечивающих его целостность. Кровеносная система имеется у немертин кольчатых червей, моллюсков, членистоногих. иглокожих, полухордовых и хордовых. Ее строение у животных разных типов сильно варьирует. Два основных продольных сосуда — спинной и брюшной, тянущиеся соответственно над и под кишечником, связаны друг с другом поперечными сосудами; последние кольцеобразно охватывают кишечник и дают ответвления к покровам и различным внутренним органам. У животных всех типов, условно объединяемых названием «беспозвоночные» кровь по брюшному сосуду течет назад, а по спинному вперед. У хордовых наоборот: по брюшному сосуду кровь течет вперед, а по спинному назад. Пульсирующей функцией, обеспечивающей движение крови, у хордовых обладает брюшной сосуд или его производное — сердце, у беспозвоночных эту функцию несет спинной сосуд. У общих предков вторичноротых животных (к ним принадлежат полухордовые, иглокожие и хордовые) кровеносная система имела очень примитивное строение, представляя собой систему лакун, т. е. полостей без эпителизированных стенок, где направление кровотока еще не установилось. В дальнейшем совершенствование кровеносной системы происходило независимо и по-разному в различных ветвях ствола вторичноротых. Поэтому состояние кровеносной системы хордовых сформировалось особым путем и не может быть выведено из развитой кровеносной системы беспозвоночных — как вторичноротых, так и (тем более) первичноротых, к которым принадлежат кольчатые черви, членистоногие, моллюски и др.
| Основные сокращения | |
| На рисунках красным цветом указана артериальная кровь, синим цветом — венозная; ак — артериальный конус; ба — брюшная аорта; бар — бедренная артерия; бв — бедренная вена; вжа — выносящая жаберная артерия; вп — воротная система почек; вс — венозный синус; всп — воротная система печени; вяв — верхняя яремная вена; ж — желудочек сердца; жа — жаберная артерия; зкв — задняя кардинальная вена; зпв — задняя полая вена; ка — кожная артерия; кв — кожная вена; кла — кожно-легочная артерия; Кп — Кювьеров проток; кр — корень аорты; ла — легочная артерия; лв — легочная вена; лда — левая дуга аорты; лж — левый желудочек; лк — луковица аорты; | лп — левое предсердие; няв — нижняя яремная вена; оса — общий ствол дуг аорты; ола — общий ствол легочных артерий; ояв — общая яремная вена; пв — подкишечная вена; пда — правая дуга аорты; пж — правый желудочек; пжа — приносящая жаберная артерия; пка — подключичная артерия; пкв — передняя кардинальная вена; пкл — подключичная вена; пп — правое предсердие; ппв — передняя полая вена; пр — предсердие; пчв — печеночная вена; поа — почечная артерия; пов — почечная вена; са — сонная артерия; сп — сонный проток; спа — спинная аорта; ха — хвостовая артерия; хв — хвостовая вена; ча — чревная артерия |
Среди хордовых наиболее примитивной кровеносной системой обладают животные подтипа бесчерепных.