Если научное и последовательное изучение процесса эмбрионального разлития обязано своим началом К. Бэру, то первыми успехами, притом весьма значительными и плодотворными, эмбриология обязана учению об эволюции, разработанному и прочно обоснованному Ч. Дарвином.
В развитии индивидуума, или онтогенезе, естественно было искать аналогий с развитием вида или более крупной группы организмов (филогенезом), и уже Дарвином были вполне использованы те эмбриологические доказательства, эволюции, которые существовали в его время. С точки зрения эволюционного учения развитие всех животных групп представляло одинаковый интерес для исследователя; оно могло выявить черты сходства строения какого-либо вида на его эмбриональных стадиях с представителями другой группы животных и дать возможность благодаря этому судить о родстве и происхождении этого вида. Можно сказать, что только благодаря эмбриологическим исследованиям удалось установить тип хордовых, к которому относятся такие несходные в окончательном своем строении представители, как оболочники, ланцетник и позвоночные; в их эмбриональном развитии всегда имеют место свойственные только им формы закладки и образования органов. Такие же характерные и постоянные способы закладки органов мы находим и в развития представителей почти каждого типа животных, как важный характерный признак этого типа.
Однако, для выяснения филогенетических вопросов, т. е. вопросов эволюции отдельных групп животных, еще большее значение, чем установление принадлежности вида к тому или другому типу, имеет возможность установить происхождение данного вида или данной группы животных, а также возможность выделить более примитивные признаки двух сходных между собой представителей. Многочисленные факты из эмбрионального развития, в особенности позвоночных животных, показали, что процесс онтогенеза и в этом отношении может дать ценные указания.
Особенно важное в этом отношении значение имеет отмеченное еще эмбриологами до-дарвиновского периода (Меккелем и Бэром) явление рекапитуляции. Рекапитуляция состоит в том, что развитие органа идет не по кратчайшему пути к той форме его, которая свойственна взрослому животному, а проходит определенные, иногда довольно сложно построенные формы, имеющие, однако, совершенно иной характер своей сложности, чем тот же орган у взрослого животного, и сходные с окончательным строением этого органа у низших представителей той же группы животных. Наиболее ярким примером рекапитуляции могут служить жаберные щели и весь принадлежащий им скелетный и кровеносный аппарат, неизменно появляющийся при эмбриональном развитии высших позвоночных, лишенных во взрослом состоянии жаберного аппарата, а также кровеносная и выделительная системы зародышей высших позвоночных, сходные ко своему строению н возникновению с таковыми рыб и амфибий.
Вопрос о связи эмбрионального развития с процессами эволюции, ставший на очередь с началом эволюционного направления в эмбриологии, был впервые разработан Ф р. Мюллером, главным образом на личиночных формах ракообразных. Эти личинки по своему строению сходны со взрослыми представителями других ракообразных, очевидно, стоящих филогенетически ниже, чем первые. Следовательно, Ф р. Мюллер имеет в виду рекапитуляцию не в отдельных органах и частях тела, а в цельном зародыше или, точнее, в цельной личинке. Отсюда он делает обобщение, что в эмбриональном развитии на более ранних стадиях формообразования проявляется строение более примитивных предков данного животного, хотя эти проявления признаков предшествующих форм оказываются гораздо менее полными, чем можно было бы ожидать, так как они сглаживаются и вторично изменяются тенденцией эмбрионального развития ускорить и упростить процесс формообразования.
Это обобщение было полностью принято Э. Геккелем, подверглось дальнейшей теоретической разработке, конкретизировано им на большом количестве примеров и получило от него наименование основного биоегенетического закона. В упрошенной формулировке закон этот выражен в положении «онтогенез повторяет собою филогенез», более же точно он сформулирован Геккелем так: «Развитие зародыша (онтогенез) есть сжатое и сокращенное повторение развития рода (филогенез), и это повторение тем полнее, чем более сохраняется вследствие постоянной наследственности первичное развитие (палингенез). Напротив, повторение это тем более неполно, чем больше произошло, вследствие изменяющегося приспособления, позднейших вторичных нарушений развития (ценогенезов). В интересах эволюционного учения задачей эмбриологов является обнаружить именно палингенезы в развитии». На основании имевшихся в то время данных по палеонтологии и сравнительной анатомии, а также на основании той оценки эмбриологических данных, которые были им сделаны и биогенетическом законе, Э. Геккель сделал попытку представить эволюционную генеалогию всех животных. Не говоря уже о том, что Геккелю при этом пришлось проявить большую изобретательность, чтобы заполнить гипотетическими формами отсутствующие в природе переходные и исходные звенья в этой генеалогии животных, все его филогенетическое дерево пришлось при дальнейших исследованиях существенно изменить, и в особенности в отношении беспозвоночных животных, суждение о филогении которых за недостатком палеонтологических сведений приходилось основывать главным образом на эмбриологических данных. При этом эмбриологические данные были использованы им очень широко, и каждой стадии, начиная от яйцевой клетки, Геккель искал соответствующую ей филогенетическую форму. Так им были созданы гипотетические стадии гастреи (соответствующей гаструле), бластеи и мореи (соответствующих бластуле и моруле); стадию гастреи он видел в типе кишечнополостных, состоящих только из эктодермы и энтодермы, а стадию бластеи — в колониальных формах жгутиконосцев (Volvox).
Мы уже говорили, что формы бластулы и еще более ранних стадий вследствие внешней пассивности бластомер являются лишь следствием физических условий среды, а не связаны с морфологией зародыша. Такие физические причины вызывают и форму Volvox, так что у нас пет никаких оснований видеть в Volvox форму, морфологически отвечающую стадии бластулы, нет также никаких оснований предполагать, что предками Metazoa были формы, сходные по строению с бластулой.
Несколько больше оснований имеется в сопоставлении гаструлы с двуслойным строением кишечнополостных, но едва ли можно схематизировать последние в гипотетическую гастрею, состоящую из индиферентной эктодермы и энтодермы, так как диференцировка клеток эктодермы является необходимым физиологическим условием существования такой двуслойной стадии.
В этой морфологической схематизации строения зародышевых стадий и в полном пренебрежении физиологической стороной; развития лежит коренная ошибка генеалогических построений Геккеля. Разделение эмбриональных признаков на палингенетические, полезные для выяснения филогении, и ценогенетические, чуждые эволюционному процессу и мешающие выяснению филогении, в известном отношении усложняло дальнейшие работы но эмбриологии и сообщало им некоторую предвзятость. Легко различить палингенез от ценогенеза в крайних их проявлениях и, конечно, например, жаберные щели зародыша высших позвоночных без труда могут быть признаны за палингенез, а образование амниона или аллантойса — за ценогенезы, но гораздо чаше зачаток оказывается одновременно и палингенетичным и ценогенетичным, и часто в самом способе ценогенизации зачатка можно найти проявление палингенеза. В тех же случаях, когда для какой-либо группы нет достаточных критериев из палеонтологии или сравнительной анатомии, ценогенез может быть ошибочно принят за палингенетическое образование. Наконец и основное отличие ценогенеза, как явления, возникающего в связи с определенными физиологическими или адаптационными потребностями зародыша, нельзя признать достаточно характерным, так как на той же физиологической почве могут возникать и палингенезы. Более того, на основании ряда примеров нужно считать, что палингенезы наблюдаются главным образом в тех органах, которые для улучшения обшей физиологии зародыша должны-начать выполнение своей специальной функции на ранних стадиях развития, а при такой ранней функциональной адаптации орган принимает строение единственно возможное на этой ранней стадии организации зародыша и это строение бывает вместе с тем сходно со строением этого органа у зародышей низших представителей данного типа животных, т. е. носит палингенетический характер.
Несомненно, что эмбриональное развитие только в том случае может дать правильное представление в частности и по филогении, если оно исследуется в целом, как определенный процесс, который состоит не только в морфологических, но и в физиологических изменениях зародыша.
Та важная роль, которую играл биогенетический закон в морфолого-эмбриологических исследованиях, сделала необходимой проверку на конкретном материале, на развитии ряда отдельных представителей, например, позвоночных, вопроса о том, в какой мере этот закон проявляется в их развитии. Эти сравнительно-эмбриологические исследования, произведенные главным образом Оппелем, Кейбелем и Менертом, показали, что в эмбриогенезе у большинства органов рост и формирование их идет с неодинаковой интенсивностью; это было уже отмечено Геккслом и названо им гетерохронией, но рассматривалось им как одно из ценогенетических отклонений онтогенеза.
Исследования (в особенности Менерта) показали, что эти гетерохронии играют настолько важную роль в развития, что ни на одной стадии весь зародыш высшего животного целиком не походит на цельное низшее животное той же группы. Если зародыш млекопитающего на определенной стадии развития имеет жаберные щели и жаберные артерии, то это не значит, что он повторяет на этой стадии строение рыбы. К этому времени другие органы, например, головной мозг, развиваясь значительно быстрее, чем у зародыша рыбы, оказывается достигшим значительно большего развития, чем у этой последней, причем развиты в типичной, характерной только для высших позвоночных форме. Из этих явлений Менерт выводит закономерность, которая состоит в том, что прогрессивные признаки и органы, т. е. важные и характерные для данного рода животных, закладываются раньше других и развиваются быстрее других; регрессивные же органы и признаки, утрачивающие свое значение для данного рода животных, закладываются в онтогенезе поздее других и развиваются замедленно.
Следовательно, в гетерохронии сказывается не только ценогенетическое отклонение, как думал Геккель, а отклонение, иногда дающее ключ к пониманию филогении данного вида животного, так как обнаружение в развитии ускорения или замедления закладки и формирования того или другого органа может показать, какие органы у данного вида являются прогрессивными и какие регрессивными. Так как вследствие гетерохроний зародыш высшего представителя не может повторить на одной и той же стадии всю организацию низшего представителя, то формулировка Геккеля для биогенетического закона во всяком случае должна быть изменена: развитие каждого органа высшего позвоночного повторяет эволюцию этого органа у низшего позвоночного.
Кроме того, так как при своей окончательной, функциональной диференцировке орган приобретает специфическое, характерное иногда даже только для данного вида, строение, то сравнение можно проводить не между вполне диференцированным органом низшего представителя и эмбриональным органом высшего, а между эмбриональными формами, между стадиями его развития у высших и у низших представителей данного типа животных.
Мы уже отмечали, что применение биогенетического закона к стадиям развития до гаструлы может повести только к ложным выводам о филогении. Но менее рискованными оказываются и попытки делать на основании этого закона выводы о происхождении и филогенетической связи видов или родов одного и того же семейства, т. е. выводы о том, какой вид надо считать первичным н исходным. Так, например, костистые рыбы на определенных и довольно поздних стадиях развития имеют непрерывный, непарный плавник, идущий но всей спине и по брюху хвостовой части тела и на заднем конце образующих хвостовой плавник в виде одной округлой лопасти. У большинства взрослых костистых рыб хвостовой плавник становится двухлопастным, но некоторые рыбы сохраняют и во взрослом состоянии круглый однолопастной плавник, соединенный со спинным и анальным плавниками. Отсюда, делают вывод, что раз у этих последних родов на всю жизнь сохраняется эмбриональная форма плавника, следовательно, они являются более первичными, даже исходными формами для родов, имеющих двулопастпой плавник. Такой вывод получается только потому, что у нас нет других проверочных критериев для суждения о большой или меньшей примитивности другого рода или вида, и он оказывается совершенно неправильным привозможности проверки иным каким-либо способом. Для иллюстрации мы возьмем эмбриональное возникновение отличий человека и антропоидных обезьян, так как истинная генеалогия их нам более известна и естественно не допускает слишком свободных выводов из данных их эмбрионального развития.
Человек отличается от человекообразных обезьян отсутствием на теле волосяного покрова, который у него сохраняется на голове, а у мужчин кроме того на верхней губе и на подбородке. У зародыша человека имеется на всем теле слабо развитый волосяной покров в виде первичного пуха, или lanugo; по такой же lanugo имеется и у зародышей человекообразных обезьян. Таким образом, первичный пух человека не отвечает дефинитивному волосяному покрову человекообразных, а так же, как первичный пух последних, отвечает сплошному волосяному покрову других млекопитающих. У зародышей шимпанзе и гориллы дефинитивный волосяной покров появляется на 5-м месяце утробной жизни в виде волос на голове, на верхней губе и на подбородке (рис. 20); только значительно позже у новорожденного детеныша 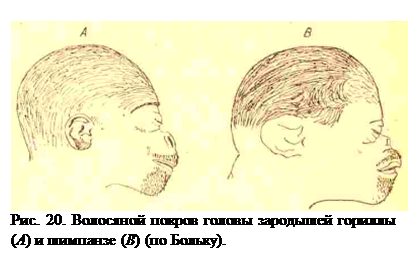 гориллы появляются волосы на всем теле. Следовательно, распределение дефинитивных полос у человека отвечает распределению у зародышей человекообразных, т. е. эмбриональное распределение полос у горилл и шимпанзе сохраняется у человека на всю жизнь. Мы должны были бы сделать вывод, что человек является более первичной формой, чем человекообразные обезьяны. Вывод совершенно неправильный, если вспомнить все основания, по которым устанавливается происхождение человека.
гориллы появляются волосы на всем теле. Следовательно, распределение дефинитивных полос у человека отвечает распределению у зародышей человекообразных, т. е. эмбриональное распределение полос у горилл и шимпанзе сохраняется у человека на всю жизнь. Мы должны были бы сделать вывод, что человек является более первичной формой, чем человекообразные обезьяны. Вывод совершенно неправильный, если вспомнить все основания, по которым устанавливается происхождение человека.
Другой пример — развитие ушной раковины у человекообразных обезьян. На одной из ранних и быстро проходящих стадий развития ушная раковина обезьян имеет округлые очертания и имеет большое количество определенно расположенных складок, а ниже ушного отверстия лишена хряща, образуя здесь мягкий круглый выступ, тождественный с мочкой (lobuhis) человека (рис. 21, А, 1). При дальнейшем развитии ушной раковины складки расправляются, и ухо разрастается в ширину, причем мочка сглаживается и исчезает; заостренная верхушка уха обезьяны развивается из особого бугорка, имеющегося на более ранней стадии и встречающегося иногда и на ушной раковине взрослого человека. Следовательно, эмбриональная форма ушной раковины обезьян сохраняется на всю жизнь в человеческой ушной раковине, из чего опять-таки исследует, что человек, есть более примитивная форма, чем обезьяна.
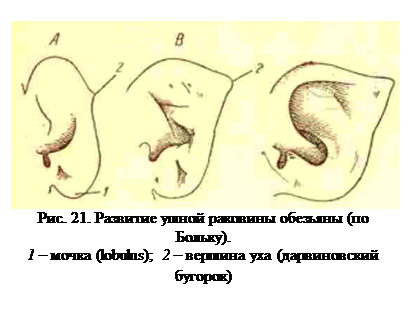 Смятая форма ушной раковины и неполный волосяной покрову человека, вероятно, являются признаками примитивности этих образований, поскольку они сходны с недоразвитым эмбриональным состоянием их у других приматов, но по этим отдельным признакам нельзя заключить, что сам человек более примитивен, так как наряду с этими признаками у него имеется ряд других, которые у обезьян представлены в более примитивной форме. Точно так же однолопастной плавник костистых рыб не говорит еще о примитивности его обладателей, так как другие органы у них могут быть более развитыми, чем у некоторых рыб с двулопастным плавником. Сохранение же у одних видов признака, являющегося только эмбриональной его формой у других, относится к той же закономерности Менерта. Волосяной покров и ушная раковина у человека стали регрессивными органами, развитие их идет медленнее, чем у обезьян, и к концу эмбрионального развития они оказываются на недоразвитой эмбриональной стадии. Это явление установлено Л. Больком, который назвал его фетализацией (от foetus — плод), и широко распространено в развитии различных животных.
Смятая форма ушной раковины и неполный волосяной покрову человека, вероятно, являются признаками примитивности этих образований, поскольку они сходны с недоразвитым эмбриональным состоянием их у других приматов, но по этим отдельным признакам нельзя заключить, что сам человек более примитивен, так как наряду с этими признаками у него имеется ряд других, которые у обезьян представлены в более примитивной форме. Точно так же однолопастной плавник костистых рыб не говорит еще о примитивности его обладателей, так как другие органы у них могут быть более развитыми, чем у некоторых рыб с двулопастным плавником. Сохранение же у одних видов признака, являющегося только эмбриональной его формой у других, относится к той же закономерности Менерта. Волосяной покров и ушная раковина у человека стали регрессивными органами, развитие их идет медленнее, чем у обезьян, и к концу эмбрионального развития они оказываются на недоразвитой эмбриональной стадии. Это явление установлено Л. Больком, который назвал его фетализацией (от foetus — плод), и широко распространено в развитии различных животных.
Таким образом, возможность применения биогенетического закона Геккеля по приведенным выше соображениям на различных стадиях развития ограничивается настолько, что он почти утратил свое значение. Явления рекапитуляции в развитии отдельных органов являются фактом, но выводы из него в духе биогенетического закона Геккеля оказываются несостоятельными. Особенно тщательному перо преисследованию были подвергнуты выводы Мюллерa и Геккеля А.Северцовым, который на основании и обширных собственных данных по эмбриологии позвоночных и появившихся до настоящего времени данных других авторов создал совершенно новую концепцию относительно связи между эмбриональным развитием и эволюцией. Северцовым на основании ряда собственных поверочных обследований развития высших и низших позвоночных не считает возможным найти в развитии высших повторения в целом организации низших; вместе с тем он приходи к выводу, что признаки крупных систематических групп возникают путем образования и суммирования отклонений в ходе эмбрионального развития.
Такие эмбриональные изменения, отражающиеся на дальнейшем строении взрослого животного, он назвал филэмбриогенезами. С этой точки зрения эмбриональные рекапитуляции перестают быть временным отражением взрослого состояния низших форм, а сами становятся материалом для эволюции. В качестве примера филэмбриогенеза можно привести то, что приблизительно на одной и той же стадии развития ящерицы (геккона) и змеи (ужа) число сомитов у последней почти вдвое больше, чем у геккона. Отсюда можно сделать вывод, что особенности развития тела змеи возникли путем появлении в развитии ящерицы отклонения, выражающегося в значительном ускорении образования сомитов (рис. 22). К филэмбриогенезам же надо отнести и замедленноеразвитие волосяного покрова и ушной раковины у человека. Филэмбриогенезы могут возникать только в период органогенеза, так как касаются не всего зародыша в целом, а только отдельных его органов.
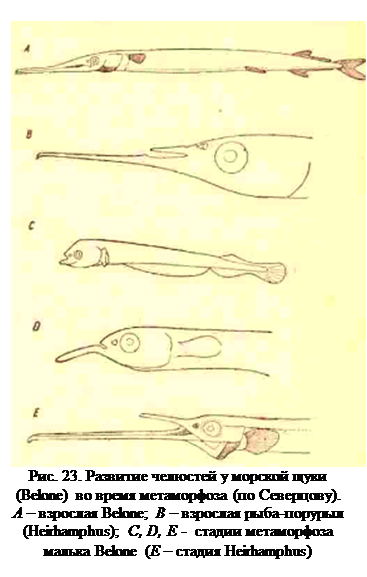
Поскольку филэмбриогенезы возникают в отдельных органах, они могут появляться и на поздних стадиях органогенеза и вызывать появление какого-либо видового или родового признака путем анаболии, т. с. путем надставок в строении более примитивно устроенного органа, типичного для других представителей этого же семейства. Так, острые и удлиненные верхняя и нижняя челюсти морской щуки Belone возникают у малька из простых укороченных челюстей, причем сначала вырастает нижняя челюсть, а позднее её догоняет в длине верхняя (рис. 23). У другого рода Heirhamphus (полурыл) того же семейства тот же процесс аноболии заканчивается на стадии длинной нижнейи верхней короткой челюсти, т. е. развитие челюстей Belone полностью рекапитулирует все стадии развития челюстей у Heirhamphus, но процесс этот идет дальше. Если филэмбриогенезы возникают в отдельных органах, то естественным был бы вывод, что организм в процессе эволюции изменяется по частям, а в таком случае непонятно, почему каждый организм имеет всегда определенную гармоничность и согласованность своих частей. Этот вопрос также хорошо разобран Северцовым и разрешается постоянно наблюдающимися в онтогенезе явлениями корреляции. Явления эти состоят в том, что изменения в одном органе всегда сопровождаются изменением в другом или даже в нескольких других органах. В одних случаях корреляция объясняется функциональной зависимостью одного органа от другого, что в грубой, но резко отчетливой форме видно в экспериментах с удалением зачатка конечности у зародыша лягушки. Если на средней стадии развития удалить у лягушки зачаток левой задней конечности, то правое полушарие среднего мозга оказывается недоразвитым; при удалении же зачатка левой конечности на более ранней стадии недоразвитым остается весь средний мозг. Соответственно сказывается на развитии среднего мозга и филэмбриогенез, выражающийся в ослаблении развития задней конечности.
 Корреляция может быть и топографической, т. е. между органами, связанными своим положением и материалом.
Корреляция может быть и топографической, т. е. между органами, связанными своим положением и материалом. 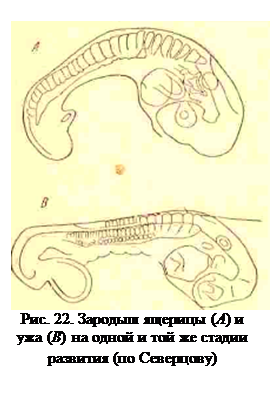 Так, при развитии части ноги лошади весь мезенхимный материал кисти пятипалогозачатка конечности уходит на образование среднего пальца; мезенхимный материал
Так, при развитии части ноги лошади весь мезенхимный материал кисти пятипалогозачатка конечности уходит на образование среднего пальца; мезенхимный материал
жаберных дуг зародыша высших позвоночных идет им образование гортанных хрящей. Возможно, что усиленное развитие мозга и черепной коробки у человека коррелятивно сопровождается уменьшением и ослаблением челюстей и зубов, а также лицевых частей по сравнению с черепом человекообразных. Во всяком случае на эмбриональное изменение одного органа зародыш реагирует изменениями в других органах, благодаря чему не только изменяются все эти органы, но и создаются новые гармонические соотношения частей. В этом сказывается особенность организма на различных стадиях своего развития и существования реагировать только как одно целое.
Значение эмбриологических данных для учения об эволюции и для выяснения филогенетических взаимоотношений между животными группами будет яснее видно на конкретных примерах, которые можно найти в описаниях развития в сравнительно-эмбриологической части книги. Классическими в этом отношении исследованиями являются упоминавшиеся во введении работы А. О. Ковалевского по развитию асцидий и ланцетника, не только по обилию и важности делаемых им выводов, но и по безусловной правильности в использовании данных онтогенеза для сопоставления признаков этих двух подтипов хордовых. Ковалевский никогда не делал каких-либо филогенетических выводов на основании стадий дробления и бластулы, как это делал Геккель для своей теории гастреи и бластеи; точно так, же мы не находим в его работах каких-либо филогенетических выводов на основании поздних стадий, когда возникают родовые и видовые признаки. Неосновательность филогенетических выводов и тю слишком ранним, и по слишком поздним стадиям развития мы постарались выяснить в этой главе.
Но эмбриональное развитие дает чрезвычайно много для суждения об эволюции животных групп на основании начального органогенеза и средних его стадий, когда, с одной стороны, проявляются морфогенетические возможности данного животного, а с другой стороны, его морфогенез носит более, общий, не затемненный частностями характер, и когда в нем проявляются только основные типовые особенности.