Живут главным образом и пресной воде или в сырой земле; поэтому они не имеют свободно плавающей личинки и выходят из яйца в виде червя. Это возможно потому, что яйца водных олигохет снабжены большим количеством желтка и кроме того помещаются по несколько штук в хитиновом коконе, содержащем кроме них еще белковую жидкость, которой зародыш питается в самом конце развития. У наземных же олигохет (дождевой червь) яйца хотя и лишены желтка, по имеют с очень ранних стадий приспособления для заглатывания окружающей их белковой жидкости. В связи с этим дробление и развитие водных и наземных олигохет обнаруживают довольно существенные различии.
 Дробление яиц водных олигохет очень неравномерно, не только потому, что неравномерно распределен желток, но и потому, что неравномерно распределена полярная протоплазма (рис. 84).
Дробление яиц водных олигохет очень неравномерно, не только потому, что неравномерно распределен желток, но и потому, что неравномерно распределена полярная протоплазма (рис. 84).
Бластомеры 2 d и 4 d в которые она попадает, гораздо крупнее других, даже обильно содержащих желток 4 А, 4 В и 4 С, и, кроме того, все бластомеры, содержащие полярную протоплазму, начинают делиться раньше других, так что синхронность дробления нарушается. К концу дробления, вследствие деления бластомер всех четырех квартетов и более медленных делений в основном квартете, последние образуют шар относительно крупных, набитых желтком клеток, анимальная половина которого покрыта, в виде колпачка слоем клеток других квартетов. Самые крупные из его клеток— производные 2 d и 4 d лежат в виде нескольких пар клеток у одного края колпачка, который является задним краем (рис. 84, D и F). Производные 2 d после нескольких делении дают 8 крупных клеток, расположенных в ряд по заднему краю колпачка, 4 справа и 4 слева от срединной линии, и называемых  эктодермальными телобластами, или эктобластами (рис. 85). Вследствие размножения клеток всего колпачка последний начинает надвигаться на шаровую поверхностькомплекса желточных клеток, и в результате края колпачка смыкаются на вегетативном полюса.
эктодермальными телобластами, или эктобластами (рис. 85). Вследствие размножения клеток всего колпачка последний начинает надвигаться на шаровую поверхностькомплекса желточных клеток, и в результате края колпачка смыкаются на вегетативном полюса.
Восемь эктодермальных телобластов во время этого процесса обрастания начинают размножаться, причем продукты их деления выдвигаются вперед от каждого телобласта правильным и длинным рядом клеток и образуют, следовательно, восемь рядов, параллельных нарастающим боковым краям колпачка и друг другу. В то же время и те две крупные задние клетки, которые, возникли из бластомеры 4 d также дают от себя по продольному ряду клеток. Они вместе с краями колпачка также смещаются к вегетативному полюсу яйца. Но, в отличие от эктотелобластов,эти два продольные ряда смещаются под край слоя эктодермы и дают здесь мезодермальные полоски. Эти клетки называются мезодермальными телобластами, или мезотелобластами, и соответствуют первичным мезодермальным клеткам.
Смыкание краев колпачка происходит не концентрически; боковые края его надвигаются к вегетативному полюсу, как два параллельных валика, тогда как сами телобласты и находящийся на противоположном им крае передний конец телобластических полосок надвигаются значительно медленнее средних их отделов; к тому времени, когда средние отделы полосок достигают вегетативного полюса, концы их доходят лишь до экватора. При окончании обрастания желточных клеток краевые ряды правой и левойсторон колпачка ложатся рядом параллельно друг к другу и образуют два срединных ряда вентральной стороны зародыша (рис. 85, А). Эти два ряда, совместно с остальными рядами, образуют парный брюшной нервный ствол, периферические нервы боковой стенки тела и кольцевую мускулатуру ее.
Мезодермальные полоски на самом переднем своем конце еще ко время обрастания желточных клеток распадаются на, мелкие, беспорядочно лежащие клетки. К концу обрастания этот передний участок каждой из полосок загибается к срединной линии, затем заворачивается концом назад, и ложится и виде рыхлой массы мезодермальных клеток параллельно остальной части полоски между нею и появляющимся здесь стомодеумом кишечника. Остальная часть каждой мезодермальной полоски после обрастания желточных клеток начинает диференцироваться на сомиты,  причем каждая клетка продольного ряда, делясь, образует такую же первичную перегородку, которая таким же путем дает сомит, как и концевые мезодермальные клетки в постларвальном отделе Polychaeta. Загнутые же передние участки полосок несколько позже вырастают в длину на протяжении нескольких передних постларвальных сомитов, вдвигаясь между этими последними и желточными клетками, и в их рыхлой массе появляются метамерные полости, постепенно уменьшающиеся к заднему, бывшему переднему, концу этого участка (рис. 86, 2). Этот участок отвечает мезодерме ларвальных сегментов, и из него впоследствии образуется мезодормальный слой с сосудами, обрастающий передний отдел кишки на протяжении так называемых головных сегментов, и временная метамерия этого участка мезодермы отвечает метамерии ларвальных сегментов.
причем каждая клетка продольного ряда, делясь, образует такую же первичную перегородку, которая таким же путем дает сомит, как и концевые мезодермальные клетки в постларвальном отделе Polychaeta. Загнутые же передние участки полосок несколько позже вырастают в длину на протяжении нескольких передних постларвальных сомитов, вдвигаясь между этими последними и желточными клетками, и в их рыхлой массе появляются метамерные полости, постепенно уменьшающиеся к заднему, бывшему переднему, концу этого участка (рис. 86, 2). Этот участок отвечает мезодерме ларвальных сегментов, и из него впоследствии образуется мезодормальный слой с сосудами, обрастающий передний отдел кишки на протяжении так называемых головных сегментов, и временная метамерия этого участка мезодермы отвечает метамерии ларвальных сегментов.
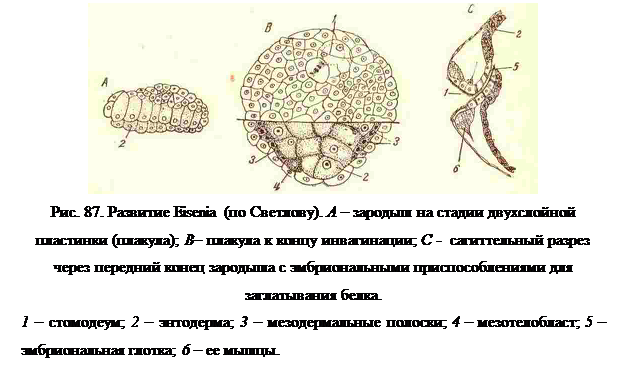 Ранние стадии развития наземных Oligoсhaeta отличаются тем, что вследствие отсутствия желтка и шара желточных клеток у них образуется вместо колпачка активных клеток выпрямленная двуслойная пластинка (плакула), в которой вместо желточных клеток имеется нижний слой этой пластинки, состоящий из некрупных энтодермальных клеток (рис. 87, А). Но дальнейшее развитие плакулы идет так, как у водных Oligoсhaeta, т. е. энтодермальная пластинка куполовидно выгибается, и начинается процесс ее обрастания верхними клетками; а так какжелточного шара нет, то этот процесс состоит в разрастании плакулы по определенной кривизне и превращении ее в замкнутый шаровидный пузырек (рис. 87, В и 89, В).
Ранние стадии развития наземных Oligoсhaeta отличаются тем, что вследствие отсутствия желтка и шара желточных клеток у них образуется вместо колпачка активных клеток выпрямленная двуслойная пластинка (плакула), в которой вместо желточных клеток имеется нижний слой этой пластинки, состоящий из некрупных энтодермальных клеток (рис. 87, А). Но дальнейшее развитие плакулы идет так, как у водных Oligoсhaeta, т. е. энтодермальная пластинка куполовидно выгибается, и начинается процесс ее обрастания верхними клетками; а так какжелточного шара нет, то этот процесс состоит в разрастании плакулы по определенной кривизне и превращении ее в замкнутый шаровидный пузырек (рис. 87, В и 89, В).
Вскоре после окончания этого процесса у зародыша диференцируется передняя часть кишечника и глотка, при помощи которой он активно заглатывает окружающий белок кокона (рис. 87, С).

 У некоторых наземных Oligoсhaeta, тело которых имеет высокое осмотическое давление, зародыш, развиваясь в дождевой влаге, постоянно поглощает воду вследствие деплазмолиза. Эта избыточная вода начинает удаляться из него с самых первых стадий развития при помощи бластомер А и В, которые специализируются на этой функции иподвергаются таким образом преждевременной диференцировке (рис. 88, А). Но тем самым они делаются уже неспособными к дальнейшему развитию и с появлением других органов осморегуляции погибают; весь зародыш, следовательно, развивается из бластомер С и D, значение которых уже заметно преобладает над А и В у всех Oligochaeta.
У некоторых наземных Oligoсhaeta, тело которых имеет высокое осмотическое давление, зародыш, развиваясь в дождевой влаге, постоянно поглощает воду вследствие деплазмолиза. Эта избыточная вода начинает удаляться из него с самых первых стадий развития при помощи бластомер А и В, которые специализируются на этой функции иподвергаются таким образом преждевременной диференцировке (рис. 88, А). Но тем самым они делаются уже неспособными к дальнейшему развитию и с появлением других органов осморегуляции погибают; весь зародыш, следовательно, развивается из бластомер С и D, значение которых уже заметно преобладает над А и В у всех Oligochaeta.
Пиявки (Hirudinea)
Про условия развития пиявок можно сказать то лее, что говорилось об Oligochaeta. Развитие только у Rhynchobdellida или хоботных пиявок (например, Clepsina) идет за счет желтка самого яйца, у остальных — за счет белка, находящегося вне яйца — в коконе. Вследствие этого развитие этих примитивных хоботных пиявок в общем идет сходно с развитием водных Oligoсhaeta, а у зародышей челюстных (Gnathobdellidae) и рыбьих пиявок (Ichthyobdellidae) при этом возникают приспособления для использования окружающего белка, которые оказывают еще более существенное влияние на развитие, чем это наблюдалось у наземных Oligochaeta. У некоторых пиявок появляется довольно хорошо развитая глотка с мускулатурой (рис. 92), иногда кишечник и тело зародыша получают способность сильно растягиваться.
Несмотря на то, что дробление у Gnathobdellidae и Ichthyobdellidae сначала в основе идет но спиральному типу, но с первых же делений этот тип искажается у них тем, чтовесь зародыш развивается главным образом из той бластомеры D стадии 4 бластомер, которая при спиральном дроблении одна даст начало имагинальным органам (рис. 90). У Gnathobdellidae микромеры 1 А, 1 В и 1 С, давпервый квартет микромер, дегенерируют, и все следующие квартеты представлены только производными D, которые дают также и всю энтодерму. Поэтому дробящееся яйцо очень рано приобретает билатеральное строение (рис. 91). А и В отделяют только клетки первого квартета, С отделяет только первый и второй квартеты. Но бластомера2 d и ее производные не могут дать сплошного эктодермального покрова, на тех ранних стадиях, когда зародыш должен начать поглощение белка и образовать для этого глотку, кишечник и эктодерму (рис. 92); последняя чрезвычайно тонка и растянута и потому является провизорной эктодермой особой формы личинок, живущей в коконе и глотающей белок. Внутри такой личинки находятся телобластические полоски клеток, состоящие из производных частью 2 d или эктотелобластов, частью же производных бластомеров 3 d, которая, подобно тому как это наблюдается у некоторых Рolychaeta становится исходной бластомерой для мезодермальных телобластов; за счет этих полосок и формируется дефинитивная эктодерма и мезодерма пиявки.
Такое же подавление специальных частей и органов трохофоры имеется и Ichthyobdеllidae (рис. 93), личинка которых, ничего общего не имеющая с трохофорой, сходна в общем с личинкой Gnathobdellidae, но ее личиночная эктодерма раздувается еще больше.
 Этими семействами пиявок завершается в ряду аннелид эволюция нового по сравнению с Рolychaeta типа развития с установкой уже не на трохофору, а на взрослого червя.
Этими семействами пиявок завершается в ряду аннелид эволюция нового по сравнению с Рolychaeta типа развития с установкой уже не на трохофору, а на взрослого червя.



Общие замечания
Относительно органогенеза аннелид вкратце отметим только следующее. Из вентральных краев первого соматобласта возникают в виде метамерных утолщений ганглии брюшной нервной цепочки, которые но мере роста каждого сегмента, остаются в виде более или менее сближенных попарно скоплений нервных клеток, соединенных в продольном направлении друг с другом пучками нервных волокон. От передней пары ганглиев вырастают волокна, соединяющие ее окологлоточной комиссурой с головным ганглием. Кпередней части головного ганглия, возникающей из клеток креста трохофоры, присоединяются с боков и сзади ганглии антенн, глаз, обонятельных ямок пальп. Антенны аннелид представляют собою плотные выступы головы, состоящие на поверхности из чувствительных клеток, а внутри сплошь заполненные нервными волокнами; пальпы же аннелид отличаются от антенн присутствием в них продолжений головной полости тела, а также и мезодермальных элементов. По сторонам каждой пары ганглиев брюшной цепочки возникает пара щетинковых мешков или параподийв виде плитных врастаний эктодермы, которые затем разрастаются в виде полых выступов, раздваивающихся на конце, на две ветви.
Соматический слой мезодермы дает продольную мускулатуру стенки тела, а спланхнический слой диференцируется на мускулатуру кишечника и его перитонеум, продуцирующий у многих форм отдельные клетки специального назначения; мезодермальные перегородки между сегментами образуют двуслойный перитонеальный диссепимент с мускулатурой между его листками. Между листками диссепимента ложатся и первичные половые клетки; из переднего листка возникает воронка метанефридия, а из заднего листка — его канал. У пиявок полости целомических мешков заполняются производными их стенок и остаются только в виде каналов, выполняющих роль кровеносных сосудов. У остальных аннелид, а частично и у пиявок кровеносные сосуды формируются также стенками целомических мешков как остатки первичной полости тела между ними. В постларвальных сегментах сначала имеется общая кровеносная полость около кишечника, но разрастанием целомических мешков она оттесняется на спинную и брюшную его стороны и в метамерные перегородки сегментов, и из нее формируются брюшной и спинной кровеносные сосуды, соединенные в перегородках кольцевыми сосудами. Кроме того, между соматоплеврой целомических мешков и эктодермой остается кожная сеть капилляров. В ларвальных сегментах сосуды не прилегают к кишечнику и образуются расхождением клеток мезодермы.
Целомические мешки возникают из 4 d. Клетки же первичной мезенхимы, производные 3-го квартета, у взрослого червя или исчезают или образуют мускулатуру глотки и диссепиментов ларвальных сегментов.