Во время роста ооцита у многих оболочников наблюдается одна особенность, состоящая в том, что некоторое количество фолликулярных клеток проникает в протоплазму ооцита и ложится у его поверхности, образуя слой так называемых тестальных клеток, или калиммоцитов. При образовании хориона последний одевает яйцо вместе c тестальными клетками. Тестальные клетки более или менее долго остаются в яйце даже во время его развития, иногда даже примешиваясь к бластомерам дробящегося яйца.
Асцидии (Ascidiae)
Одиночные асциди (Monascidiae).Эмбриональное развитие. Яйцо выходит в воду, одетое фолликулярными клетками, которые выделяют хорион, а сами затем приобретают пенистое строение и поддерживают яйцо у поверхности воды. Однако, у некоторых асцидий яйца большую или меньшую часть своего развития проходят в клоакальной или перибранхиальной полости матери и не образуют пенистой оболочки. Такие яйца обычно более богаты желтком.
Детально прослеженное на Styela (Cynthia) формирование и развитие яиц, бедных желтком, представляет вместе с тем один из лучших примеров развития с весьма стойкой предварительной структурой протоплазмы, о возникновении которой мы уже говорили в общей части (стр. 17). Желток в ооците Styela состоит из более грубых зерен в вегетативном полушарии и мелкой зернистости в анимальном, а весь поверхностный слой ооцита содержит желтый пигмент. При редукционном делении крупный зародышевый пузырек, или ядро ооцита, теряет оболочку и ядерный сок и выходит в протоплазму анимального полушария, а желтая протоплазма отходит в вегетативное полушарие (рис. 8). При вхождении сперматозоида желтая протоплазма снова смещается и образует серповидную фигуру, занимающую половину экватора яйца, а под ней, в виде второго серпа, располагается прозрачная протоплазма из анимального полушария; однако, после слияния пронуклеусов прозрачная протоплазма уходит с поверхности вглубь яйца, расходясь по анимальyому полушарию. Перед первым делением яйца грубозернистая протоплазма вегетативного полушария разделяется по виду на два сорта: один, занимающий большую часть вегетативного полушария, содержит более густо сконцентрированный темносерый желток и мало протоплазмы; другой, занимающий в виде серпа половину экватора, не занятую желтым серпом, имеет более светлую окраску, содержит больше протоплазмы и меньше желтка. Плоскость первого деления яйца разделяети светлосерый и желтый серп посередине, и каждая из этих двух бластомер получает все еорта протоплазмы яйца (рис. 8, С). Как показывает дальнейшее развитие, эта плоскость соответствует сагиттальной медиальной плоскости тела зародыша и личинки, причем желтый серп отвечает задней стороне тела личинки, а светлосерый серп — передней стороне его.
|
|
Если придать яйцу такое положение, сохраняя которое оно даст зародыша в нормальном положении, т. е. спинной стороной кверху, брюшной книзу, то анимальyый полюс яйца должен быть направлен вперед и вниз, а вегетативный — назад и вверх; главная ось яйца должна быть наклонена под углом в 45° к горизонту. Второе деление отделяет две более крупные передние бластомеры от двух меньших задних. В каждую из них попадает и прозрачная и грубозернистая протоплазма, ио светлосерая протоплазма попадает только в две передние бластомеры, а желтая — только в две задние клетки. Третье деление проходит экваториально (по отношению к анимальному и вегетативному полюсам), причем плоскость его проходит так, что две анимальные передние бластомеры меньше двух передних вегетативных, а две задние анимальные бластомеры больше двух задних вегетативных; кроме того, передние вегетативные и задние аиималъ- ные бластомеры вытянуты в поперечном направлении, а передние анимальные н задние вегетативные — в продольном. Таким образом, на стадии 8 бластомер уже достаточно ясно намечается билатеральный тип дробления яйца асцидий. На последующих стадиях дробления билатеральиость его выражается в том, что вытянутые в поперечном направлении бластомсры делятся продольно, а вытянутые в продольном направлении при том же делении делятся в поперечном направлении. Например, при четвертом делении передние анимальные и задние вегетативные бластомеры делятся поперечно и дают поперечно вытянутые бластомеры следующей стадии, а передние вегетативные и задние анимальные делятся продольно и дают вытянутые продольные 4 бластомеры стадии 16 бластомер.
|
|
При пятом делении определяется судьба 4 аннмальных бластомер, которые состоят из прозрачной протоплазмы и дают только наружные покровы личинки. Вегетативные бластомеры при третьем и четвертом делениях состоят: передние из темно- серой и светлосерой протоплазмы, две задние — из темносерой и желтой протоплазмы, а другие две задние бластомеры — почти из одной только желтой протоплазмы. При пятом же делении происходит окончательное распределение желтой, темносерой н светлосерой протоплазмы по отдельным бластомерам. Самые передние светлосерые бластомеры дают хорду и нервную пластинку, второй ряд и две средние бластомеры третьего ряда из темиосерой протоплазмы дают кишечник, а задний полукруг желтых бластомер дает всю мезодерму. Другими словами, светлосерый передний серн яйца дает материал хорды и нервной системы, а желтый серп — материал мезодермы.
|
|
При шестом делении (на 64 бластомеры) четыре светлосерые бластомеры делятся в поперечном направлении и образуют две дугообразные группы: из них передняя дуга представляет собою материал будущей нервной системы, а позади нее лежащая дуга — материал будущей хордальной пластинки (стр.348, А). Бластомеры боковых участков желтой задней дуги делятся поперечным делением на наружные клетки — материал будущей мускулатуры и внутренние — материал мезенхимы, каковыми являются и все остальные желтые бластомсры, прилегающие к материалу кишечника.
Седьмое деление происходит с большим ускорением деления бластомер, дающих мезодерму, хорду и нервную систему, и с запаздыванием деления эктодермальных (анимальных) бластомер, так что получается стадия 76 клеток; а затем делятся анимальные клетки, давая стадию 112 бластомер, а не 128, так как материал кишечника не делится. При этом вегетативное полушарие значительно сплющивается благодаря начинающейся гаструляции. Последняя состоит в постепенном погружении всех вегетативных клеток в бластоцель, вследствие чего вегетативная поверхность вдавливается. При этом сначала вдвигаются внутрь средние клетки полушария, т. е. материал кишечника, а за ними следуют клетки мезенхимы и хорды, так что по краям образовавшегося таким образом широкого овального бластопора располагаются по переднему полукругу клетки зачатка нервной пластинки, но сторонам бластопора — клетки мускульного зачатка и назади — не ушедшие в бластоцель клетки мезенхимы.
Бластопор из овального становится Т-образным вследствие надвигания на него боковых краев с клетками мышечного зачатка, которые начинают погружаться в бластоцель, так что к боковым краям бластопора приближаются теперь клетки эктодермыПередняя часть бластопора более круто и глубоко опускается в гастроцель, чем задняя. Затем дуга нервного зачатка, состоящая из четырех рядов клеток, начинает сильно надвигаться в виде крышки на бластопор спереди, частью путем размножения клеток, частью вследствие того, что боковые части этого подковообразного зачатка начинают сдвигаться к срединной линии, так что зачаток принимает форму четыреугольной пластинки. Однако, в передней части нервного зачатка смещения клеток к срединной линии ие происходит, благодаря чему эта часть остается более широкой и сохраняет дугообразную форму. Так же удлиняется и суживается хордальный зачаток, подстилающий нервную пластинку снизу. Вследствие этого бластопор уменьшается и остается открытым только назади.
К концу гаструляции зародыш имеет строение мешка, состоящего из двух слоев, которые мы, не зная образования деталей зачатков, назвали бы эктодермой и энтодермой. Зная же происхождение зачатков, мы должны считать, что в состав наружного эктодермального слоя входят клетки двоякого происхождения: клетки наружного покрова и клетки нервной пластинки, возникающей из одного зачатка с клетками хорды; последние, однако, входят в состав внутреннего, «эктодермального» слоя. Внутренний же слой состоит не только из энтодермы, т. е. зачатка кишечника, но и из материала хорды и мезодермы.
Во второй период гаструляции, т. е. когда происходит надвигание на отверстие бластопора его переднего края или передней губы, материал хорды вытягивается назад и приходит в соприкосновение с вдвигающимся ему навстречу материалом мезодермы, образуя вместе с ней и с эктодермальным материалом внутренний слой гаструлы. Подобный тесный контакт хордального материала с материалом нервной пластинки и мезодермы является необходимым условием для дальнейшего развития и организации мезодермы. Если отделить в яйце асцидии на стадии 4 бластомер две передние от двух задних бластомер, то передние дают самодифереицнровкои хорду и нервную систему, но из двух задних бластомер образуется беспорядочный комок клеток, скоро прекращающий свое развитие, так как здесь нет хордального материала. Так как энтодермалъный материал в нормальной гаструле асцидий отделен от хорды мезодермальными клетками, то кишечник запаздывает в своем окончательном оформлении и диференцировке и не функционирует у личинки асцидий.
Во второй половине гаструляции клетки эктодермы, нервной пластинки и хорды принимают эпителиальный характер, клетки же кишечника и мезодермы сохраняют рыхлое расположение, хотя и лежат вполне симметрично.
По окончании гаструляции нервная пластинка начинает опускаться ниже прилегающей с боков эктодермы и сгибается в виде желобка, который смыкается краями в нервную трубку и покрывается вместе с тем сверху слоем эктодермы. Эктодерма вместе с верхней финкой нервной трубки надвигается на бластопор, лежащий у ее заднего края в виде очень небольшого отверстия, и отверстие его сохраняется, но открывается не прямо наружу, а в задний конец просвета нервной трубки. Таким образом, канал нервной трубки переходит назади в бластопор, а через него в полость кишечника, вследствие чего бластопор получает теперь название нервно-кишечного канала. Замыкание нервной трубки начинается с заднего конца от области бластопора. Отчасти по этой причине, отчасти вследствие того, что передняя часть нервной пластинки шире остальной, самый, передний конец нервной трубки долгое время остается открытым наружу небольшим отверстием, называемым невропором, а передний конец самой нервной трубки образует крупное мозговое расширение. Нервно-кишечный канал ограничеи спереди клетками дна нервной трубки и подстилающей ее хорды, с боков — клетками мышечного зачатка и сзади — клетками зачатка кишечника. Зачаток же мезенхимы, как начавший раньше мигрировать внутрь гаструлы, оказывается лежащим у переднего конца мышечного зачатка, но сторонам кишечного зачатка, и не соприкасается с нервно-кишечным каналом.
 Затем кишечный зачаток, представленный до сих пор только вентральной стенкой гастрального впячения, смыкается своими краями под хордальным зачатком в мешок. В то же время начинается рост заднего конца зародыша благодаря размножению всех клеток, примыкающих к нервно-кишечному каналу, включая также и эктодерму. Вследствие этого зародыш становится грушевидным, суженный конец его увеличивается в длину н загибается на вентральную сторону в виде довольно длинного цилиндрического продолжения тела, представляющего собой зачаток хвоста личинки.
Затем кишечный зачаток, представленный до сих пор только вентральной стенкой гастрального впячения, смыкается своими краями под хордальным зачатком в мешок. В то же время начинается рост заднего конца зародыша благодаря размножению всех клеток, примыкающих к нервно-кишечному каналу, включая также и эктодерму. Вследствие этого зародыш становится грушевидным, суженный конец его увеличивается в длину н загибается на вентральную сторону в виде довольно длинного цилиндрического продолжения тела, представляющего собой зачаток хвоста личинки.
Из сказанного следует, что вырастают в длину и входят в состав хвоста зародышазктодерма, узкое продолжение нервной трубки, боковые группы мускульных клеток,зачаток хорды и задний участок вентральной стенки кишечника (рис. 238). Последняявытягивается в виде клеточного тяжа, называемого хвостовой кишкой. Хордальныйзачаток вытягивается также в однородный клеточный тяж, дисковидные клетки которого плотно прижаты друг к другу. С появлением хвостовой кишки сообщение полостикишечника с полостью нервной трубки, или нервно-кишечный канал, исчезает. С окончанием роста заднего конца эктодермахвоста образует оторочку хвостовогоплавника, на переднем конце тела образуются прикрепительные папиллы, ипроисходит диференцировка мышечныхклеток и всего нервного зачатка. Зародыш превращается в хвостатую личинку  асцидии.
асцидии.
Хвост личинки еще больше удлиняется, но не размножением клеток, а гистологическими изменениями зачатков,сопровождающими функциональную диференцировку. Клетки хордального зачатка вырабатывают цилиндрический стержень вязкого вещества, причем самиклетки хорды остаются на поверхностистержня, сильно распластываясь на неми образуя его клеточную оболочку. Вместе с тем хорда смещается несколько назад и лежит теперь только в пределах хвоста. Мышечные клетки диференцируются в продольные мышечные тяжи, и этот зачаток также удлиняется.
В туловищном отделе, на дорзальной его стороне впереди невропора, появляется стомодеальное впячение эктодермы, но оно прорывается в кишечник только в конце личиночной жизни, анальное же отверстие появляется только после метаморфоза, так что личинка не может питаться. Невропор закрывается. Мозговое расширение имеет вид полого мешка шаровидной формы (рис. 239, 5). На внутренней стороне его венеральной стенки диференцируется простого строения статолит, состоящий из одной только клетки (11), которая тонкой ножкой соединена со стенкой мозгового расширения (5): полость этого расширения играет роль статоциста. На внутренней же стороне задней стенкимозгового расширения возникает пигментированный бокаловидный глаз со стекловидным телом (6). Мезенхима лежит в задней части туловищного отдела и еще не диференцируется; только часть мезенхимы выходит через покровный эпителий наружу и выделяет особую тунику, одевающую личинку.
За время личиночной жизни организация туловищного отдела заметно усложняется. На дорзальной стороне тела по сторонам нервной трубки образуются два мешковидные впячения эктодермы с узким входным отверстием; это атриальные впячения, или парный зачаток перибранхиадьной полости (рис. 239, 12), охватывающей с боков заднюю часть кишечного мешка; последний дает позднее два выступа, прорывающихся в перибранхиальной полости отверстиями — первой парой жаберных щелей.
На заднем конце кишечного мешка с правой стороны вырастает отросток, который сперва направляется назад, затем дугообразно заворачивает вперед к левой стороне кишечного мешка. Вершина, образуемая этим выростом дуги, вздувается в виде зачатка желудка, нисходящая узкая часть выроста становится пищеводом, который затем соединяется со стомодеумом, а восходящая узкая часть — задней кишкой, заканчивающейся слепо (рис. 239, 7 и 10). Сам же обширный кишечный мешок представляет собой жаберный отдел кишечника, или жаберную глотку. Стенки его, как и некоторые другие ткани, далее к моменту метаморфоза остаются наполненными желтком. На передней стенке жаберной глотки узкий медиальный участок се длфереицнруется в полоску мерцательных клеток и одноклеточных железок, вдавленную в стенку глотки в виде желобка, который представляет собою эндостиль.
К концу личиночной жизни стенка стомодеума дает узкое впячение с мерцательными ресничками внутри, направленное к нижней стороне мозгового мешка. Последний дает навстречу выступу стомодеума свой выступ, соединяющийся с первым. Это двойное образование представляет собой гипофиз.
Метаморфоз. Период свободно плавающей личинки длится у асцидий недолго и, как у всех сидячих животных, имеет значение главным образом для расселения потомства. Вскоре личинка опускается на дно и прикрепляется своими тремя прикрепительными папиллами или сосочками к субстрату, так что хвост ее торчит вертикально кверху (рис. 240). К этому моменту нерибранхиальиые полости разрастаются, прикрывая с боков значительную часть поверхности жаберной глотки, а число жаберных щелей увеличивается до 6—8 пар. Слепо заканчивающаяся у личинки задняя кишка прорывается в левую перибранхиальную полость анальным отверстием. Сразу вслед за прикреплением начинается процесс превращения, носящий характер регрессивного или дегенеративного метаморфоза. Хвост быстро укорачивается, а находящиеся в нем хорда, мускулатура, нервный спинной тяж и хвостовая кишка распадаются на отдельные клетки, поедаемые фагоцитами, которые возникают из начавших дифсренцироваться клеток мезенхимы. В туловищном отделе мозговой мешок также редуцируется, но здесь редукция личиночных органов сопровождается возникновением имагинальных. Трубка гипофиза, образуемая стомодеумом, уничтожается, но часть гипофиза, происходящая из стенки мозгового мешка, не только остается, но и дает размножением своих клеток весь имагинальный нервный узелок.
Участок стенки тела между группой прикрепительных сосочков, лежащей на переднем конце личинки, и ротовым отверстием ее, лежащим на спинной стороне, начинает расти значительно быстрее других участков, вследствие чего ротовое отверстие все дальше отодвигается от места прикрепления и все больше приближается к заднему концу личинки, который после прикрепления становится верхним концом асцидии. Таким образом, рот описывает дугу почти в 180°, и вместе с ним поворачивается на 180° и весь комплекс внутренних органов (рис. 240), в котором в это время происходят свои изменения.
Во время всего метаморфоза появление имагинальных органов происходит в общем следующим образом. Участок эктодермы, где открываются наружу обе перибранхиальные полости, или карманы, втягивается вглубь тела, а отверстия перибраихнальных полостей постепенно расширяются. Таким образом, оба кармана сообщаются теперь с наружной  средой не непосредственно, а через обширную клоакальную полость, а сами становятся как бы продолжениями ее, охватывающими со всех сторон широкий жаберный отдел кишечника. В эту же клоакальную полость открывается теперь и анальное отверстие. Жаберный отдел кишечника потребляет в это время весь оставшийся в его клетках желток. Количество жаберных щелей быстро увеличивается; кроме того каждая из них разделяется растущим от дорзальной стороны язычком на две, так же как это имеет место у кишечнодышащихВ приведенной выше картине эмбрионального развития мы не находим никакихпризнаков энтероцельной закладки мезодермы, свойственной всем описанным выше Deuterostomia,и никаких целомическихмешков. Причина этоговыясняется при изучении возникновения новых мезодермальных образованийпри метаморфозе. Дело втом, что у зародыша образуются только мышцы и мезенхима, и о совершеннонет перитонеума, и кишечник лежит в первичнойполости тела. Образованиеперитонеальных зачатков начинается только во время метаморфоза в виде закладки перикардия сердца и пары эпикардиев. Перикардий возникает каквыступ вентральной стенкижаберного отдела кишки взадней ее части около входав пищевод (рис. 241, В, 2).Он отшнуровывается откишки в виде замкнутогомешочка, после чего надорзальной стенке его, обращенной к кишке, образуется входящая в мешочекскладка (рис. 241, С, 2 и 3). Она глубоко вдается в перикардий, и листки ее расходятся, образуя вторую полость, куда проникают жидкость первичной полости тела иклетки мезенхимы. Этот внутренний мешочек перикардия становится пульсирующимсердцем.
средой не непосредственно, а через обширную клоакальную полость, а сами становятся как бы продолжениями ее, охватывающими со всех сторон широкий жаберный отдел кишечника. В эту же клоакальную полость открывается теперь и анальное отверстие. Жаберный отдел кишечника потребляет в это время весь оставшийся в его клетках желток. Количество жаберных щелей быстро увеличивается; кроме того каждая из них разделяется растущим от дорзальной стороны язычком на две, так же как это имеет место у кишечнодышащихВ приведенной выше картине эмбрионального развития мы не находим никакихпризнаков энтероцельной закладки мезодермы, свойственной всем описанным выше Deuterostomia,и никаких целомическихмешков. Причина этоговыясняется при изучении возникновения новых мезодермальных образованийпри метаморфозе. Дело втом, что у зародыша образуются только мышцы и мезенхима, и о совершеннонет перитонеума, и кишечник лежит в первичнойполости тела. Образованиеперитонеальных зачатков начинается только во время метаморфоза в виде закладки перикардия сердца и пары эпикардиев. Перикардий возникает каквыступ вентральной стенкижаберного отдела кишки взадней ее части около входав пищевод (рис. 241, В, 2).Он отшнуровывается откишки в виде замкнутогомешочка, после чего надорзальной стенке его, обращенной к кишке, образуется входящая в мешочекскладка (рис. 241, С, 2 и 3). Она глубоко вдается в перикардий, и листки ее расходятся, образуя вторую полость, куда проникают жидкость первичной полости тела иклетки мезенхимы. Этот внутренний мешочек перикардия становится пульсирующимсердцем.
Несколько выше зачатка перикардия от стенки жаберного отдела обособляются ещедва полых выступа, направленных назад к желудку; это правый и левый эпикардии.Отделившись от кишечника в виде удлиненных тонкостенных мешков, оба эпикардияначинают расширяться и вытеснять первичную полость тела; в конце концов они соприкасаются друг с другом и образуют мезентерий кишечника.
Таким образом, у зародыша асцидии и у личинки закладывается только часть мезодермальиых образований, свойственных асцидиям. Так как мышцы у них являютсячисто личиночным органом, то возникновение их у зародыша нужно рассматривать какпреждевременную диференцировку с полным уничтожением этого органа при метаморфозе. Такую же преждевременную диференцировку претерпевают и клетки мезенхимы, участвующей у личинки в образовании туники.Но мезенхима при метаморфозе не уничтожается,а передиференцируется в мезенхиму асцидии.Позже всего возникает перитонеум целомическихмешков; вероятно по этой причине дающая емуначало стенка жаберного отдела кишки дометаморфоза содержит в своих клетках желток,нужный для образования этих запоздалых зачатков мезодермы.
 Эпикардий, однако, имеет и другое значение,особенно в жизни сложных асцидий, так как у них при почковании он дает ряд органов новыхособей. Возможно, что эта роль эпикардия является следствием того же запоздалого его образования, благодаря которому его клетки являются у асцидий наиболее молодыми мезодермальными элементами.
Эпикардий, однако, имеет и другое значение,особенно в жизни сложных асцидий, так как у них при почковании он дает ряд органов новыхособей. Возможно, что эта роль эпикардия является следствием того же запоздалого его образования, благодаря которому его клетки являются у асцидий наиболее молодыми мезодермальными элементами.
У некоторых асцидий с относительно большимколичеством желтка в яйце свободная личиночнаястадия выпадает, точнее говоря, все стадии личинки и начало регрессивного метаморфоза проходят у них в яйцевой оболочке. Вылупляющееся животное поэтому уже неспособно к плаванию; оно падает на дно и прикрепляется.
У асцидий все стадии до личинки очень мало способны к регуляции; у личинки регулятивные способности ограничиваются некоторыми исправлениями повреждений прикрепительных сосочков. Эта ограниченность регуляции отчасти связана с постоянством числа клеток на определенных стадиях зародыша и у личинки. С наступлением метаморфоза связанность перемещения клеток при регуляторных процессах, обусловленная предварительной структурой яйца, исчезает, и многие асцидии во взрослом состоянии обладают в высокой степени способностью к регенерации и бесполому размножению.
Сложные асцидии (Synaseidiae). Большинство асцидий образуют колонии почкованием или делением исходной особи, развивающейся из личинки. Очень распространено также бесполое размножение столонами.
Деление колониальных, или сложных, асцидий происходит сильным вырастанием в длину нижней части асцидии, которая оказывается вследствие этого сидящей на стебле. Вместе со стеблем растут и два эпикардия, тянущиеся вдоль всего стебля. Эпикардии расчленяются, начиная от нижнего конца, образованием поперечных перехватов. Стебель отделяется от тела асцидии и, соответственно разделившимся участкам эпикардпен, распадается на отдельные новые особи (рис. 242). При таком делении, насколько известно, все внутренние органы возникают из эпикардия. В таких колониях особи остаются соединенными между собой только общей туникой.
 Почкование (например, у Botrillus) происходит путем появления на теле материнскойособи наружного выпячивания эктодермы, в который входит выступ перибранхиальнойполости. Новые особи возникают, следовательно, целиком из эктодермальныхпроизводных (эпикардий в почкованиине участвует); особи колонии связаныдруг с другом и общей туникой, и отростками от нижних частей тела.
Почкование (например, у Botrillus) происходит путем появления на теле материнскойособи наружного выпячивания эктодермы, в который входит выступ перибранхиальнойполости. Новые особи возникают, следовательно, целиком из эктодермальныхпроизводных (эпикардий в почкованиине участвует); особи колонии связаныдруг с другом и общей туникой, и отростками от нижних частей тела.
Размножение столонами состоит вобразовании выроста от нижнего концаособи, развившейся из яйца и называемой оозооидом, или от того же конца ранее отпочковавшейся особи; этот вырост,называемый столоном, имеет вид трубки,внутрь которой врастает пластинчатыйотросток эпикардия. На столоне образуются боковые веточки, которые вздуваются и образуют новые особи. Последние или остаются соединенными со столоном (Сlavellina) или отрываются от него и сохраняют связь как особи однойколонии, только благодаря общей тунике(Distaplia). У Distaplia и некоторых других сложных асцидий яйцо содержитмного желтка, личинка выходит из негозначительно продвинутой в своем развитии, и на теле ее имеются уже первыедве почки.
 Пиросомы (Lucida). Каждая особьпелагической колонии пиросомы образуеттолько одно яйцо, одетое фолликулом;Последний, отделив к яйцу большое количество так называемых тестальныхклеток, принимает вид кубического эпителия, который участвует в переработкепитательных веществ тела матери в желток яйца и растягивается по мере увеличения объема яйца. Фолликулярныйэпителий остается на яйце, при его оплодотворении и во время его развития, котороетакже сопровождается ростом зародыша и растягиванием фолликула. Все развитиеяйца проходит сначала в яйцеводе, а затем в клоакальной и правой перибранхиальной полостях.
Пиросомы (Lucida). Каждая особьпелагической колонии пиросомы образуеттолько одно яйцо, одетое фолликулом;Последний, отделив к яйцу большое количество так называемых тестальныхклеток, принимает вид кубического эпителия, который участвует в переработкепитательных веществ тела матери в желток яйца и растягивается по мере увеличения объема яйца. Фолликулярныйэпителий остается на яйце, при его оплодотворении и во время его развития, котороетакже сопровождается ростом зародыша и растягиванием фолликула. Все развитиеяйца проходит сначала в яйцеводе, а затем в клоакальной и правой перибранхиальной полостях.
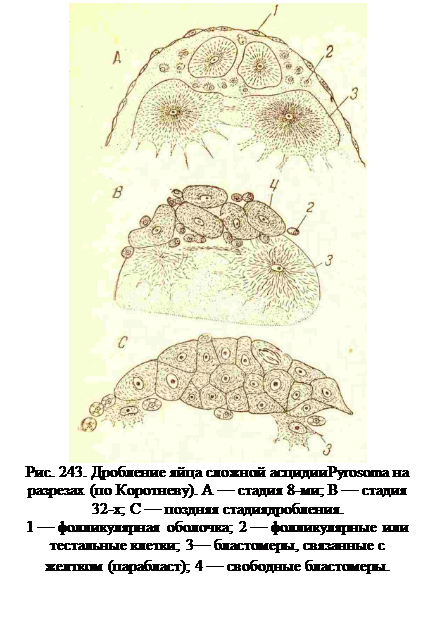
 Как было указано выше, личинки асцидий не могут питаться, и поэтому у тех асцидий, почкование которых начинается в личинке или в начале метаморфоза, это размножение возможно потому, что яйца их содержат количество желтка, достаточное не толькодля полного развития личинки, но и для образования первых почек. У пелагических колониальных асцидий, или пиросом (род Pyrosoma), из яйца образуются одновременно и исходнаяособь, или оозооид, и еще четырепроизошедших от них особи, илибластозооиды. Количество желткав яйце у них очень велико, так что протоплазма лежит в виде небольшого скопления на анимальном полюсе большой желточной массы, и дробление идет дискоидально (рис. 243 и 244). Послеоплодотворения зародышевый дискприподнимается в виде бугорка,и так же приподняты бластомерыпервых делении (рис. 243).
Как было указано выше, личинки асцидий не могут питаться, и поэтому у тех асцидий, почкование которых начинается в личинке или в начале метаморфоза, это размножение возможно потому, что яйца их содержат количество желтка, достаточное не толькодля полного развития личинки, но и для образования первых почек. У пелагических колониальных асцидий, или пиросом (род Pyrosoma), из яйца образуются одновременно и исходнаяособь, или оозооид, и еще четырепроизошедших от них особи, илибластозооиды. Количество желткав яйце у них очень велико, так что протоплазма лежит в виде небольшого скопления на анимальном полюсе большой желточной массы, и дробление идет дискоидально (рис. 243 и 244). Послеоплодотворения зародышевый дискприподнимается в виде бугорка,и так же приподняты бластомерыпервых делении (рис. 243).
Четыре первых борозды дробления проходят меридианально,перпендикулярно к поверхности яйца, по уже со стадии 8 бластомер средние 4 бластомеры (бластоциты) отделяются от поверхности яйца, а 4 краевые (мероциты) остаются в соединении с желтком (рис. 243). При следующих делениях мероциты ложатся вровень с поверхностью желтка, бластоциты образуют круглую пластинку не связанных с желтком клеток.
Тестальные (фолликулярные) клетки попадают под оболочку яйца в довольно большом количестве и примешиваются к бластомерам во время ранних стадий дробления(рис. 243, 2 и 244). На более поздних стадиях их количество уменьшается; во всяком случае, они не участвуют в образовании зародыша.
 Количество мероцитов увеличивается врезультате деления, и они образуют своейпротоплазмой хорошо заметный синцитиальный слой на участке поверхности желтка,прикрытом бластодиском. Они перерабатывают желток и являются поэтому желточнойэнтодермой, которая не принимает участия вформировании зародыша и исчезает к концупотребления желтка. Кишечная энтодермавозникает путем подворачивания одногоиз краев бластодиска (заднего) под наружный слой клеток в виде второго слоя, который растет вперед и подстилает весь бластодиск (рис. 245). Край бластодиска, где происходит подворачивание энтодермы, очерченрезче других краев его и представляет собоюбластопор, вернее, верхнюю губу его.
Количество мероцитов увеличивается врезультате деления, и они образуют своейпротоплазмой хорошо заметный синцитиальный слой на участке поверхности желтка,прикрытом бластодиском. Они перерабатывают желток и являются поэтому желточнойэнтодермой, которая не принимает участия вформировании зародыша и исчезает к концупотребления желтка. Кишечная энтодермавозникает путем подворачивания одногоиз краев бластодиска (заднего) под наружный слой клеток в виде второго слоя, который растет вперед и подстилает весь бластодиск (рис. 245). Край бластодиска, где происходит подворачивание энтодермы, очерченрезче других краев его и представляет собоюбластопор, вернее, верхнюю губу его.
Эти ранние стадии развития пиросомпоразительно похожи на дробление и гаструляцию также, развивающегося из дискоидально дробящегося яйца костистых рыб.Нижний листок этой двуслойной стадии на периферии распадается на клетки, представляющие собою мезенхиму, а центральначасть нижнего листка, приподнимаясь наджелтком, дает зачаток жаберного отдела кишечника и рядом с ним, повидимому, зачаток перикардия. Вопрос о возникновении зачатков других органов недостаточно выяснен. Нервная система появляется в виде впячения эктодермы, впячиванием же эктодермывозникают и две трубкообразные периб ранхнальные полости. Таким образом, возникаеторганизация первой особи, которая имеет жаберный отдел кишечника, перикардий,нервную систему и перибранхиальные мешки (рис. 246. D). Желтка в яйце в это времяеще очень много, и потому органы первой особи, или оозооида, распластаны по его поверхности и представляют собою далеко еще не закончившие своего развития зачатки, на не настоящие органы. Из них несомненно функционирует только сердце с перикардием, которое интенсивно пульсирует. Оозооид занимает центральную часть бластодиска, края которого постепенно надвигаются на желток, но к стадии оозооида еще не покрывают всего желтка.
Далее начинается рост оозооида, конец которого, противоположный по положениюотверстию перибранхиальных полостей, начинает выдвигаться в виде язычка, свободнонависающего над поверхностью желтка, н бластодиска; язычок этот содержит в себе перибранхиальные трубки,кишечник с эндостилем и перикардий,причем все эти органы образуются в нем прямым разрастанием соответствующих органов оозооида (рис. 247).Получается своеобразный короткийстолон, на котором четырьмя перехватами, появляющимися одновременно на всем его протяжении, отделяется от оозооида 4 бластозооида(рис. 247, D). Первый служит для бластозооидов питающей особью, иликормилкой, так как его сердце гонит в них вместе с кровью питательныевещества из желтка, перерабатываемого мероцитами оозооида.
Только с появлением 4 бластозооидов заканчивается процесс обрастания желтка бластодиском. Потребление желтка вместе е тем ускоряетсяи заканчивается поглощением мероцитами всего желтка. Одновременно с израсходованием желтка гибнетоозооид, но сердце продолжает битьсядаже в уже распадающемся оозооиде.
 4 первых бластозооида, соединенные друг с другом в цепочку, вырабатывают каждый органы, свойственныеэтому животному. Эти первые особи колонии затем ложатся в виде розетки, и из этой молодой колонии путем почкования образуются все особивзрослой колонии.
4 первых бластозооида, соединенные друг с другом в цепочку, вырабатывают каждый органы, свойственныеэтому животному. Эти первые особи колонии затем ложатся в виде розетки, и из этой молодой колонии путем почкования образуются все особивзрослой колонии.
Эти первые бластозооиды, развивающиеся за счет желтка, перерабатываемого для них мероцитами оозооида, появляются одновременно все четыре, и вначале представляют собой как бы едппын организм, который потом разделяется. В дальнейшем почки возникают в известной последовательности одна за другой, и каждая из них с момента появления имеет отдельную, индивидуальную организацию. Постоянной остается при этом и группировка всех вновь отпочковывающихся особен, что обуславливает как образование определенной формы Pyrosoma в целом, так и особый характер взаимной связи отдельных ее особей.
Так как пиросомы в виде колонии ведут пелагический образ жизни, то стадия специальной пелагической личинки, необходимая для распространения сидячих асцидий, у них отсутствует. Поэтому и все развитие пиросом утрачивает свойственную другим асцидиям установку на личинку, т. е. в нем отсутствуют зачатки хорды, хвостовой мускулатуры и длинной нервной трубки с нервно-кишечным каналом.