То, что V-области кодируются более чем одним генным сегментом, установлено в 1978 г., когда был закончен первый анализ нуклеотидной последовательности одного из генов иммуноглобулинов-гена Vλ, выделенного из эмбриона мыши. Вместо того чтобы кодировать все 110 аминокислот Vλ-области, ген Vλ кодировал только 97 N-концевых аминокислот. Оказалось, что оставшиеся 13 аминокислот этой области кодируются отдельным сегментом ДНК, расположенным на сотни тысяч нуклеотидов дальше и получившим название соединительного или Jλ-сегмента (не путать с боковой J-цепью, которая кодируется в другой части генома). Jλ-сегмент всегда примыкает к Сλ,-гену, от которого он отделен интроном. В ходе развития В-клетки Vλ-ген (который мы будем далее называть Vλ-сегментом), поскольку он кодирует лишь часть Vλ-области полипептидной цепи, транслоцируется таким образом, что оказывается рядом с Jλ-сегментом, так что в ДНК получается последовательность Vλ-Jλ-интрон-Сλ. Эта последовательность транскрибируется в молекулы РНК, из которых интроны после удаляются, так что после сплайсинга образуются молекулы мРНК с примыкающими друг к другу последовательностями V, J и С. Именно эти молекулы мРНК транслируются в полипептиды легких цепей.
Впоследствии было установлено, что у мышей имеется несколько разных J-сегментов в каждом пуле генов для иммуноглобулинов: по одному ассоциировано с каждым С-геном в генном пуле λ-цепей и по четыре – в генных пулах х- и Н-цепей (каждый С-ген отделен от соседнего с ним J-сегмента интроном). В период развития В-клеток в генных пулах х- и Н-цепей любой V-сегмент может объединиться с любым J-сегментом, что увеличивает в четыре раза число V-областей, которые эти пулы могут производить. Кроме того, существуют некоторые различия в точном месте соединения V-J, и это приводит к еще большему разнообразию аминокислотных последовательностей. Существенно то, что сайт соединения V-J кодирует часть третьей гипервариабельной области легкой цепи.
После соединения V и J все «лишние» J-сегменты «ниже» присоединенного J-сегмента транскрибируются, а затем их последовательности, так же как и последовательности интронов, вырезаются из РНК-транскрипта (рис. 6). Появляется все больше данных в пользу того, что соединение тех или иных V- и J-сегментов ДНК в процессе развития В-лимфоцитов сопровождается элиминацией всей лежащей между ними ДНК (см. рис. 6). Конкретный механизм соединения сегментов, которые могут быть удалены друг от друга на сотни тысяч нуклеотидов, остается загадкой. Однако обнаружены специфические, весьма консервативные последовательности ДНК, примыкающие «снизу» к V-сегментам и «сверху» к J-сегментам.
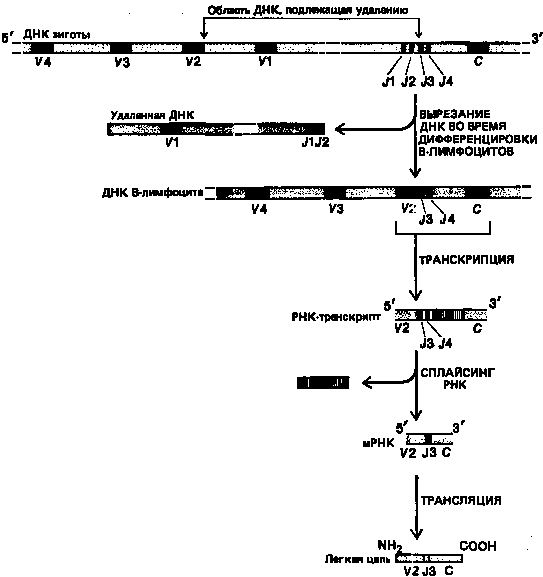
Рисунок 6. Процесс соединения V-J, с которым связано образование х-легкой цепи у мыши.
Предполагается, что они могут служить сайтами узнавания для ферментов сайт-специфической комбинации ДНК (рис. 7), но такого рода ферменты пока не идентифированы.
V -область каждой Н-цепи кодируют три генных сегмента
Сборка гена, кодирующего вариабельную область тяжелой (Vh) цепи процесс еще более сложный, чем сборка гена вариабельной легкой цепи (Vl). Здесь тоже участвует J-сегмент, однако в дополнение к нему некоторые аминокислоты третьего гипервариабельного участка V-области кодируются еще одним генным сегментом, получившим название D-сегмента (diversity gene segment). Число различных D-сегментов неизвестно, но у мыши их не менее десятка. В результате серии сайт-специфических перекомбинаций D-сегмент соединяется с каким-либо из Vн или с каким-либо из Jн-сегментов, что приводит к созданию функционального Vн-гена. Существование отдельных D-сегментов еще по меньшей мере в 10 раз увеличивает число различных Vн-областей, которые могут образоваться у мыши.

Рисунок 7. Схема, иллюстрирующая возможную роль специфических последовательностей ДНК (показаны цветными), расположенных «ниже» V-сегмента и «выше» J-сегмента, в соединении V-J. Взаимодействие между этими последовательностями, вероятно, происходит при участии сайт-специфической рекомбинационной системы, катализирующей разрыв и воссоединение двойных спиралей ДНК в области этих последовательностей.
Разнообразие антител увеличивается благодаря соматической рекомбинации, соединению легких и тяжелых цепей в различных сочетаниях и возникновению соматических мутаций. В процессе эволюции иммунной системы выработался целый ряд различных механизмов, приводящих к большому разнообразию антиген-связывающих участков антител. Только часть из этих механизмов связана с описан выше соматическими перестройками ДНК в ходе развития В-лимфоцитов. Эксперименты по подсчету числа генов с использованием метода гибридизации ДНК показывают, что в геноме мыши, видимо, содержится несколько сотен Vх-сегментов, сходное число Vн-сегментов и два Vλ-сегмента. Из этого можно вычислить, что путем комбинированы личных унаследованных V-, D- и J-сегментов у мыши может образоваться по меньшей мере 10000 разных Vн-областей и 1000 разных VL-областей.
Один простой, но весьма важный механизм, сильно увеличивающий разнообразие антител, состоит в комбинировании различных L- и Н-цепей. Поскольку в антиген-связывающий участок антитела вносят вклад как L-, так и Н-цепи, у животного с 1000 генов, кодирующих Vl-o6ласти, и с 10000 генов, кодирующих Vн-области, продукты этих генов могли бы образовать 1000 х 1000 различных комбинаций, т.е. 107 разных антиген-связывающих участков, если предположить, что при этом любая L-цепь может сочетаться с любой Н-цепью.
Недавно было показано, что в генах V-области и поблизости от них происходят соматические мутации; вероятно, это увеличивает число разных антител по крайней мере в 10-100 раз. Механизмы, благодаря которым мутации возникают именно здесь, неизвестны. Однако было обнаружено, что мутации встречаются гораздо чаще в антителах IgG и IgA, чем в кодируемых Vн-геном антителах IgM. Это, пожалуй, неудивительно, так как молекулы IgM вырабатываются на ранних стадиях иммунного ответа, a IgG и IgА появляются относительно поздно. Поэтому В-клетки, переключившиеся на IgG или IgA, как правило, претерпели большее число делений, чем В-клетки, продолжающие вырабатывать IgM, и поэтому, скорее всего, накопили больше мутаций. С другой стороны, может существовать механизм, повышающий частоту мутирования V-генов после того, как произойдет переключение с IgM на иммуноглобулины других классов. Как бы было, изменение антиген-связывающих участков в результате соматических мутаций может вести к тому, что антиген будет особенно эффективно вызывать пролиферацию тех В-клеток, у которых измененные антитела приобрели высокое сродство к антигену; возможно, что именно этим (по крайней мере отчасти) объясняется повышение сродства антител, наблюдаемое после иммунизации (созревание сродства). Таким образом, соматические мутации могут не только увеличивать разнообразие антител, но и служить для тонкой подстройки антител при иммунном ответе.