В-лимфоциты. Введение
В-лимфоциты - функциональный тип лимфоцитов, играющих важную роль в обеспечении гуморального иммунитета. При контакте с антигеном или стимуляции со стороны T-клеток B-лимфоциты трансформируются в плазматические клетки, способные к продукции антител всех классов. Активированные B-лимфоциты могут превращаться в B-клетки памяти для обеспечения в будущем более быстрого иммунного ответа на АГ. Помимо продукции АТ, В-клетки могут:
· Быть АПК
· Продуцировать цитокины и экзосомы
Субпопуляции В-лимфоцитов
Под субпопуляциями понимают разновидности клеток определенного типа, характеризующиеся наличием устойчивых различий по функциям и связанным с ними молекулярным маркерам. Выделяют несколько субпопуляций В-клеток (В-эффекторов):
· В1 - локализуются в брюшной и плевральной полости. Секретируют антитела, а также выявляют в селезенке, где на их долю приходится 1–5% от числа всех В-клеток. Могут мигрировать через сальник в слизистую оболочку кишечника и брыжеечные л/у. В л/у у мыши отсутствуют. Экспрессируют мембранную молекулу СD5 (один из маркеров Т-клеток). Для В1-клеток характерен «активированный фенотип», что проявляется в экспрессии на их поверхности костимулирующих молекул СD80 и СD86. Это свойство обеспечивает способность В1-лимфоцитов выполнять функции АПК. В эмбриональном периоде В1-клетки мигрируют в серозные полости, где они существуют в течение всей жизни организма. Способны к самоподдержанию путем очень медленной пролиферации, восполняющей убыль клеток, погибающих по механизму апоптоза. Подразделяются на две дополнительные субпопуляции:
1. В1а-клетки. Несут на поверхности молекулы IgM и CD5. Экспрессируют BCR, содержащий мембранную форму IgM. В1а-клетки в онтогенезе появляются раньше других субпопуляций — еще до рождения. Они развиваются в печени плода при участии IL-5 и IL-10 из клеток-предшественников, отличных от таковых у обычных В-клеток.
|
|
2. В1b-клетки. Экспрессируют BCR, содержащий мембранную форму IgM. Развиваются в печени эмбрионов, а после рождения — в костном мозгу из других клеток-предшественников. Расселяются на периферии несколько позже В1а-клеток — непосредственно перед рождением и после него. Могут мигрировать в серозные полости и сохраняются там путем самоподдержания.
· В2 – основная популяция (обычные В-клетки). Зрелые экспрессируют низкое количество мембранного IgM и высокое количество IgD. Локализуются преимущественно в селезенке, ККМ, л/у, пейеровых бляшках и отдельных фолликулах лимфоидной ткани кишечника. В2-клетки составляют подавляющее большинство циркулирующих В-лимфоцитов и играют основную роль в гуморальном иммунном ответе. В2-лимфоциты рециркулируют значительно слабее, чем Т-клетки. Вне фолликулов В2-клетки живут около недели, но в естественном микроокружении существуют в течение нескольких недель и даже месяцев. Срок полуобновления при действии повреждающих факторов составляет 13 сут.
· В клетки маргинальной зоны (MZB). Локализуются в маргинальной зоне селезенки, отделяющей белую пульпу от красной. Фенотипически сходны с В2 и происходят от тех же костно-мозговых клеток-предшественников. Основной мембранный иммуноглобулин IgM и экспрессируется сильнее, чем на В2-клетках. IgD так же присутствует на мембране, но, в очень малом количестве. Сходны фенотипически с активированными В-лимфоцитами. Имеют молекулы CD69, CD25, CD38, и в малом количестве CD23. Имеется молекула CD1d, участвующая в презентации липидных антигенов. Срок жизни MZB-лимфоцитов сопоставим со сроком жизни организма. Участвуют в гуморальном иммунном ответе на возбудители, поступающие в кровоток. Осуществляют тимуснезависимый иммунный ответ на инкапсулированные патогены. Благодаря сильной экспрессии молекул MHC-II и костимулирующих молекул обладают выраженной способностью к взаимодействию с Т-хелперами. В этих клетках не происходит переключения классов иммуноглобулинов.
|
|
· В-хелперы
· В-памяти
· В-супрессоры
· В-регуляторные
В-лимфоциты экспрессируют иммуноглобулиновый рецептор на своей поверхности для распознавания антигенов — BCR (содержится около 150 000 комплексов BCR). IgM-антитела, секретируемые клеткой, являются пентамерами (5 мономерных структур IgM), мембранный IgM В-клеточного рецептора представляет собой мономер. После активации антигеном класс антигенраспознающего рецептора В-клетки может изменяться вместо IgM и IgD на IgG, IgE и IgA. Н-цепи мембранных иммуноглобулинов отличаются от растворимых 2-мя С-концевыми участками — трансмембранного и цитоплазматического. Трансмембранный встраивается в мембрану клетки.
Взаимодействие с МНС
Важную роль играют В-пимфоциты в процессах формирования гуморального иммунного ответа. Наряду с дендритными клетками и макрофагами В-лимфоциты входят в семейство антигенпредставляющих клеток (АПК). Только эти клетки экспрессируют на мембране MHC-I и МНСII. Распознавание АП-рецепторами Т-лимфоцитов комплекса МНС-II-антиген является важнейшим этапом в индукции процесса образования антитед В-лимфоцитами. Все остальные клетки организма несут на мембране MHC-I (локализуются на всех ядросодержащих клетках организма, а не только на специализированных АПК), функциями АПК не обладают. При взаимодействии BCR с антигеном образующийся комплекс ВСR-антиген пиноцитируется В-лимфоцитом и внутри клетки фрагментируется с помощью внутриклеточных ферментов. В процессе разрушения антигена его пептидные фрагменты встраиваются в молекулы МНС-II и экспрессируются на поверхности В-клетки. Этот этап именуется представдением антигена, а кдетки, представляющие антиген Т-лимфоцитам, называют АПК.
|
|
Рецепторный аппарат
П. Эрлих предсказал существование рецепторов на поверхности клеток, а Ф.Бернет основал клонально-селекционную теорию. Во 2-й половине 60-х годов ХХ века рецепторы В-клеток были идентифицированы как мембранные иммуноглобулины (mIg). Позже было показано, что BCR (B-cell receptor) содержит, помимо иммуноглобулина, дополнительные белковые молекулы, обеспечивающие передачу сигнала внутрь клетки.
Мембранный иммуноглобулин — специфический маркер В-клеток, поскольку он
На поверхности зрелых В-клеток экспрессирован специфический маркёр – мембранный иммуноглобулин, который отсутствует на других клетках + присутствует ещё IgD. На наивных (не активированных) В-клетках начиная со стадии незрелых имеется мембранный иммуноглобулин класса M (IgM). Число молекул иммуноглобулинов на поверхности наивной В-клетки составляет около 150 000. В процессе иммунного ответа происходит переключение классов иммуноглобулинов на:
· IgG (несут клетки преимущественно в крови и вторичных л/о
· IgA (несут клетки слизистых оболочек)
· IgЕ
Поверхностные иммуноглобулины построены из идентичных Н- и L-цепей и имеют один и тот же изотип и аллотип, а также обладают одинаковой специфичностью к антигенам и идентичны по идиотипу. Разные В-клетки отличаются по специфичности мембранных рецепторов. Клетки, происходящие от одного предшественника (клоны В-клеток), имеют идентичные мембранные иммуноглобулины. Все клоны В-клеток формируют антигенраспознающий репертуар В-лимфоцитов, который обеспечивает распознавание всех возможных конфигураций молекул (эпитопов)
Структура мембранных иммуноглобулинов идентична структуре растворимых антител кроме С-концевого участка. Мембранный иммуноглобулин содержит гидрофобный трансмембранный участок, обеспечивающий заякоривание молекулы на клеточной мембране. Цитоплазматический участок мембранных иммуноглобулинов очень короткий.
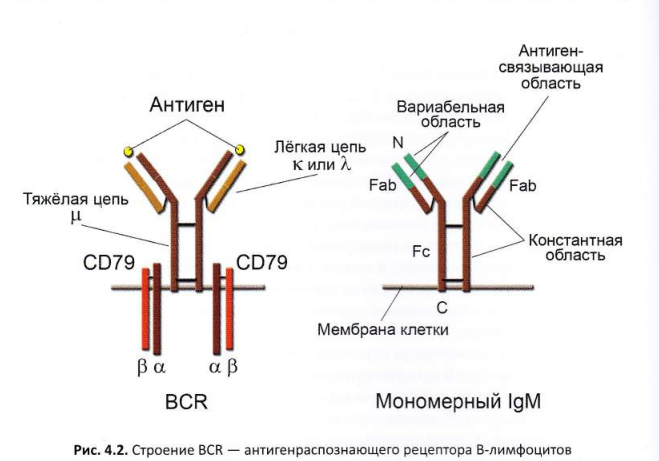
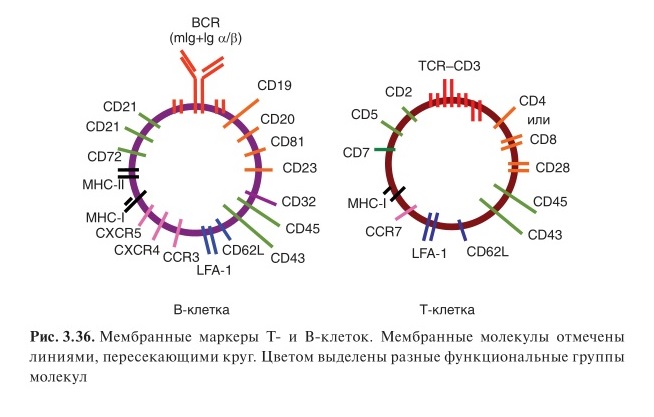
В состав BCR помимо иммуноглобулина входит:
· СD79a и СD79b — составляют интегральную чаcть BCR. Варианты a и b называют еще Igα и Igβ. Участвуют в передаче сигнала
· CD19, CD21 и CD81 — функционально ассоциированы с ним и формируют физическую связь с BCR только при активации клетки.
1. CD19 относят к суперсемейству иммуноглобулинов, играет важную сигнальную функцию. Около цитоплазматической части молекулы CD19 располагается липидная киназа PI 3K и обеспечивает запуск, передачу активационных сигналов при связывании антигена. Определяется на В-клетках на стадии про-В.
2. CD21 — рецептор для компонентов комплемента (CR2), участвующий в усилении антигенного сигнала и регуляции активности В-лимфоцитов.
3. СD81 (тетраспанин). Функция этой молекулы точно не определена, но вероятно участвует в передаче активационного сигнала.
Молекулы, связанные с BCR, рассматривают как маркеры В-лимфоцитов и их экспрессию определяют (с помощью моноклональных антител) для подсчета численности В-клеток. Строгоспецифичной для В-лимфоцитов является только молекула CD19. Маркером фолликулярных, В2-клеток является мембранная молекула CD23.
На поверхности В-лимфоцитов под влиянием активации экспрессируются другие молекулы для выполнения функций:
· В-клетки несут на поверхности молекулы MHC не только I, но и II класса
· Костимулирующие молекулы CD40, CD86, а при активации — также CD80.
Благодаря экспрессии этих молекул В-лимфоциты выполняют роль АПК. В-клетки экспрессируют молекулы адгезии, позволяющие им мигрировать из сосудов и перемещаться в тканях:
· β 1 -интегрины VLA-2 и VLA-4
· β 2 -интегрин LFA-1
· L-селектин CD62L
Присутствие на их поверхности Fc-рецепторов (FcγRIIВ — CD32) и рецепторов для комплемента (СR2) в регуляции активности В-клеток играет болльшую роль, чем для выполнения ими эффекторных функций.
В-клетки экспрессируют важнейшие рецепторы для цитокинов:
· IL-4-6
· IL-1-2
· IL-10
На поверхности так же присутствуют рецепторы для цитокинов семейства TNF, защищающие В-клетки от развития апоптоза и выполняют гомеостатическую функцию, поддерживая численность этих клеток на постоянном уровне. Главные факторы — цитокины BAFF и APRIL:
· BAFF (B-cell activating factor of TNF family) — BAFF-R, BCMS, TAC-1,
· APRIL (A proliferation inducing ligand) — HSPG
На В-лимфоцитах представлены рецепторы для хемокинов:
· CXCR4 (для SDF-1)
· CXCR5 (для BLC, служащего основным хемоаттрактантом для наивных В-клеток)
· CCR3 (для эотаксинов)
· CCR6 (для LARC)
В-клетки различных субпопуляций присутствуют в:
· межфолликулярных областях
· в мозговых шнурах лимфатических узлов
· краевых зонах белой пульпы селезенки
· а также в дерме, собственной пластине слизистых оболочек, подслизистом слое.
У взрослого человека и большинства млекопитающих, включая грызунов, В-лимфоциты развиваются в ККМ, а в период эмбрионального развития в фетальной печени. У птиц В-клетки развиваются в бурсе Фабриция, у жвачных млекопитающих — в кишечнике. Заключительные этапы развития В-клетки проходят вне костного мозга во вторичных лимфоидных органах.
В-клетки развиваются из общего лимфоидного предшественника, который происходит от полипотентной кроветворной стволовой клетки. Ранние родоначальные клетки, а также общий миелоидный предшественник лишены маркеров, свойственных основным линиям кроветворных клеток. Подобно стволовым клеткам, экспрессируют:
· молекулу CD34
· несут мембранную молекулу CD45RA
Обычно лимфоидные предшественники имеют трецепторы для гемопоэтических факторов:
· c-Kit (для SCF)
· Flt-3 (для лимфопоэтического фактора Flt-3L)
Все это отличает их от общих миелоидных предшественников. Часть лимфоидных предшественников несет на своей поверхности:
· Либо молекулу CD7
· Либо CD10
· Далее со временем экспрессируется рецептор для IL-7 (CD127) — фактора роста для всех незрелых форм лимфоидных клеток. Они дают начало В-лимфоцитам и, частично, плазмоцитоидным дендритным клеткам
Корецепторный аппарат
Для активации В-лимфоцита существенно-значимой является экспрессия на его мембране корецепmорньнх молекул с сигнальной функцией, включающих:
· CD19
· CD21 (рецептор CR2 для компонентов комплемента iС3b, C3d и C3dg). Определяется на стадии незредых В-лимфоцитов.
· ТАРА- 1 (CD81). Участвует во взаимодействии В-клетки с Т-лимфоцитом.