Вопрос о «болевых» рецепторах является одним из традиционно сложных и дискуссионных в проблеме боли и аналгезии. Несмотря на все возрастающее количество исследований, посвященных начальным, периферическим механизмам возникновения боли, до настоящего времени нет четких представлений ни о морфофункциональной организации аппарата, воспринимающего болевые воздействия, ни о существовании самих воздействий, адекватных для болевых рецепторов. Общим свойством разных no своей модальности стимулов, способных вызвать боль, является повреждение тканей — ноцицепция, по Ч. Шеррингтону. Поэтому в современной литературе широко используется термин «ноцицептор» в качестве семантического аналога «болевого рецептора», хотя в смысловом отношении они, как и понятия «боль» и «ноцицепция», не эквивалентны.
Существует немало различных классификаций ноцицепторов — по модальности воздействия, по способности отвечать на определенный или различные ноцицептивные стимулы, по принадлежности к тонким миелиновым и безмиелиновым волокнам, по локализации в различных тканях и т.д. Однако такое разделение, в основе которого лежат разные подходы и принципы, не вносит большой ясности в понимание природы рецептора и не дает ответа на основной традиционный вопрос, специфичны ли болевые рецепторы или боль является функцией интенсивности и синхронности любого воздействия.
М. Фрей, по-видимому, первым выдвинул гипотезу о существовании специфических болевых рецепторов и предположил, что эти рецепторы представлены свободными неинкапсулированными нервными окончаниями. Согласно современным данным, они в большом количестве содержатся в различных тканях и органах и имеют множество концевых разветвлений с мелкими аксоплазматическими отростками, которые и являются структурами, активируемыми болевым воздействием.
Было показано, что при раздражении кожи человека через металлические электроды, прикосновение которых практически не ощущалось, выявлялись «точки», пороговая стимуляция которых воспринималась как резкая нестерпимая боль. Представление о специфических ноцицепторах, как о свободных немиелинизированных нервных окончаниях, получило дальнейшее развитие и экспериментальное подтверждение. Кроме того, в коже и, особенно, в дентине зубов были обнаружены своеобразные комплексы свободных нервных окончаний с клетками иннервируемой ткани, которые рассматриваются как сложные рецепторы болевой чувствительности.
Отличительной особенностью многих свободных немиелинизированных окончаний является их высокая хемочувствительность. Допускается даже, что болевой рецептор по своей природе не ноцицептивен, а «хемоцептивен», поскольку любое воздействие, приводящее к повреждению тканей и являющееся адекватным для ноцицептора, сопровождается высвобождением алгогенных химических агентов. Выделяют 3 типа таких веществ — тканевые, плазменные и выделяющиеся из нервных окончаний. Предложено немало гипотез о ноцицептивных механизмах алгогенных субстанций. Допускается, что субстанции, содержащиеся в основном в тканях, непосредственно активируют концевые разветвления немиелинизированных волокон и вызывают импульсную активность в высокопороговых кожных, висцеральных и мышечных афферентах. Они вызывают ощущение боли у человека и псевдоаффективную ноцицептивную реакцию у животных при аппликации на ткани. Другие, сами не вызывающие боли, усиливают эффект ноцицептивного воздействия иной модальности. Третьи выделяются непосредственно из терминалей и взаимодействуют с рецепторами, локализованными на их мембране, и, деполяризуя ее, вызывают генерацию импульсного ноцицептивного потока. Предполагается, что субстанция Р, содержащаяся в сенсорных нейронах спинномозговых ганглиев, действует и как синаптический передатчик в нейронах заднего рога спинного мозга. В качестве химических агентов, активирующих свободные нервные окончания, рассматриваются не идентифицированные до конца вещества или продукты разрушения тканей, образующиеся при сильных повреждающих воздействиях, при воспалении, при локальной гипоксии. Эндогенно образующиеся алгогенные вещества играют существенную роль в возникновении мышечной и висцеральной боли. Кроме того, эти вещества в субноцицептивных концентрациях способны активировать и полимодальные ноцицепторы. С выделением эндогенных алгоиндукторных субстанций связывают явление сенситизации при воспалении и разномодальных повреждениях кожи.
Свободные нервные окончания активируются и интенсивным механическим воздействием, вызывающим их деформацию, обусловленную сжатием ткани, растяжением полого органа с одновременным сокращением его гладкой мускулатуры. Эти окончания могут выполнять функцию специфических висцеральных ноницепторов в миокарде, в легких, в желчных протоках. Они активировались только такой интенсивностью стимулов, которая вызывала псевдоаффективные ответные реакции.
Одновременно с представлениями о специфичности болевых рецепторов существуют и активно обсуждаются несколько разновидностей теории интенсивности А. Гольдшайдера, согласно которым боль возникает не в результате раздражения специальных «ноцицепторов», а вследствие избыточной активации всех типов рецепторов различных сенсорных модальностей, которые в норме реагируют только на неболевые, «неноцицептивные» стимулы. В формировании боли в этом случае главенствующее значение имеет интенсивность воздействия, закодированная в характере импульсной активности в афферентных проводниках, а также пространственно-временное соотношение афферентной информации, конвергенция и суммация афферентных потоков в ЦНС. Не вдаваясь в детальную дискуссию о доказательствах или опровержениях различных разновидностей этой теории, следует отметить, что в последние годы получены весьма убедительные данные о наличии «неспецифических ноцицепторов» в сердце, кишечнике, легких и их важной роли в возникновении висцеральной боли.
В настоящее время считается общепризнанным, что основными проводниками кожной и висцеральной болевой чувствительности являются тонкие миелиновые А-δ- и безмиелиновые С-волокна, различающиеся по ряду физиологических свойств. Более того, допускается, что сами «ноцицепторы» представлены концевыми разветвлениями этих волокон в тканях.
По-видимому, впервые Я. Цоттерман предположил, что именно с активацией С-волокон связано ощущение боли. Это предположение в последующем было подтверждено рядом исследований, но оно по-прежнему основывалось на сравнении ощущений человека с данными, полученными при регистрации активности С-афферентов у животных. У кошек и приматов неоднократно показано, что различное ноцицептивное раздражение кожи вызывает импульсную активность в С-афферентах, причем часть из них активировалась всеми тремя видами ноцицептивных стимулов, а часть — только механическим и термическим раздражениями. Характерно, что частота и длительность импульсной активности в С-волокнах отчетливо коррелировала с интенсивностью и продолжительностью ноцицептивного воздействия. Предполагается, что у некоторых видов животных С-афференты имеют ведущее значение в генезе термической боли.
Только с помощью микронейрографического метода регистрации активности отдельных С-волокон у человека было доказано, что ощущение боли возникает тогда, когда активируются эти афференты. Установлено, что у человека С-полимодальные волокна активируются нагреванием кожи в пределах +40... +49 °С, уколом, щипком, накожной аппликацией или внутрикожным введением гистамина, калия хлорида и других алгогенных субстанций. Рецептивные кожные поля С-афферентов имели площадь от 1 мм2 до 1 см2. Порог их термического ноцицептивного раздражения составлял +41...+47 °С, что хорошо соответствует порогу ощущения термической боли. У большинства С-афферентов число и частота импульсов линейно возрастали с увеличением интенсивности термического раздражения. Прямая зависимость была выявлена между характером импульсной активности и субъективной оценкой болевого ощущения. При длительном ноцицептивном воздействии в С-афферентах наблюдался феномен утомления с одновременным уменьшением боли, но через непродолжительное время возникала сенситизация, проявляющаяся резким снижением порога термического раздражения С-волокон, и возрастала интенсивность болевого ощущения. Весьма характерно, что одиночное раздражение афферентных нервов у человека интенсивностью, достаточной для активации С-волокон, вызывало ощущение умеренной плохо локализуемой боли, однако при аналогичной по силе, но более высокочастотной стимуляции уже через 3 с боль воспринималась как очень сильная и даже нестерпимая. Эти данные свидетельствуют, что временная суммация импульсных потоков, приходящих в головной мозг по С-афферентам, играет чрезвычайно важную роль в формировании боли.
А-δ-афференты тоже активируются механическим и термическим ноцицептивными раздражепиями кожи. Предполагается, что с импульсной активностью в бимодальных А-δ-волокнах связано то начальное ощущение острой колющей боли, которое испытывает человек сразу же после сильного термического по вреждения кожи - локального ожога, прикосновения к небольшому горячему предмету. Считается, что А-δ-волокна играют незначительную роль в передаче ноцицептивной информации из внутренних органов за исключением легких, где они связаны с ирритантными рецепторами.
С активацией определенного калибра афферентных волокон связывают различные типы болевых ощущений: так называемую первичную — светлую, коротколатентную, хорошо локализованную и качественно детерминированную боль, и вторичную — темную, длиннолатенгную, плохо локализованную, тягостную, тупую боль. Такое разделение боли, многократно установленное в различных психофизиологических исследованиях при разных ноцицептивных воздействиях, в настоящее время является общепринятым. При сопоставлении ощущения боли у человека со спектром вовлеченных в возбуждение афферентных волокон при раздражении кожных нервов было убедительно показано, что «первичная» боль связана с афферентной импульсацией в А-δ-волокнах, а «вторичная» — с С-волокнами. Нарушение проведения возбуждения в миелиновых волокнах приводит к ослаблению «первичной» боли, но «вторичная» боль может даже усиливаться.
Однако весь комплекс представленных данных ни в коей мере не должен инициировать представление об А-δ- и С-волокнах, как об однородной группе проводников исключительно болевых сигналов. Широко используемые в литературе термины «болевые», «ноцицептивные» афференты конечно нельзя понимать буквально. Они являются полимодальными сенсорными системами. Эти волокна передают информацию, не связанную с ноцицепцией, и активируются неповреждающими термическими и механическими воздействиями. Более того, активация небольшой группы С-волокон недостаточна для возникновения ощущения боли у человека, но боль может быть вызвана высокочастотной стимуляцией низкопороговых толстых миелиновых афферентов группы А-а.
Независимо от того, каковы механизмы возникновения ноцицептивной информации на периферии, в формировании боли ключевое значение имеют процессы, происходящие в ЦНС. Именно на основе центральных механизмов: конвергенции, суммации, взаимодействия быстрой миелинизированной и медленной немиелинизированной систем на разных уровнях головного мозга — создаются ощущение и качественная окраска боли при действии различных ноцицептивных раздражении.
Первым центральным звеном, воспринимающим разномодальную афферентную информацию, является нейрональная система заднего рога спинного мозга. По современным данным, она представляет собой цитоархитектонически весьма сложную структуру, а в функциональном отношении рассматривается как своеобразный первичный интегративный центр сенсорной информации. Именно здесь происходит взаимодействие между каналами болевой и неболевой чувствительности, на основании которого формируется поток восходящей импульсации нового качества. В настоящее время в мировой литературе имеется огромное количество публикаций, посвященных морфофункциональной организации этой области спинного мозга и ее роли в переработке, формировании и передаче ноцицептивной информации, которые обобщены в ряде монографий и обзоров.

Рис 1 - Распределение окончаний первичных афферентов в спинном мозге
Слева - окончания афферентных волокон разных типов от сухожильных рецепторов, мышечных веретен и кожи, не связанных с ноцицепцией; справа - окончания безмиелиноных С афферентов и тонких миелиновых волокон, связанных с ноцицепцией, I — VI — слои серого вещества спинного мозга по В. Rexed.
Распределение первичных афферентов в заднем роге спинного мозга. Афферентные волокна различного калибра по-разному распределяются уже в задних корешках перед их входом в спинной мозг — толстые миелинизированные проводники занимают их медиальную часть, а тонкие миелинизированные и С-волокна располагаются более латерально. Перерезка этих латеральных пучков сопровождается уменьшением боли. После входа в спинной мозг А-δ- и С-волокна в составе тракта Лиссауэра идут в каудальном и ростральном направлениях в пределах 1 — 2 сегментов и оканчиваются в дорсальной части заднего рога. Имеется определенное различие в окончании А-δ-, С-волокон, передающих ноцицептивную информацию, и А-δ-волокон, связанных с механорецепторами.
Установлено, что в I пластине, содержащей крупные, маргинальные клетки, оканчиваются А-δ-волокна, активируемые термическим и механическим ноцицептивными раздражениями, и С-полимодальные ноцицентивные афференты. Широко ветвящиеся тонкие терминали С-волокон образуют синапсы на дендритах маргинальных клеток и аксоаксональные контакты на терминалях других афферентных волокон. Желатинозная субстанция является основным местом окончания С-волокон, а также других афферентов, которые входят в нее с дорсальной поверхности, и, широко разветвляясь, диффузно распределяются во всей ее толще, достигая вентральной границы. А-δ-волокпа также оканчиваются в желатинозной субстанции. Причем А-δ-афференты, связанные с механорецепторами, оканчиваются в центральной части II пластины и в III пластине и более вентрально, тогда как ноцицентивные А-δ-волокна распределяются в верхних отделах II пластины.
Толстые афферентные волокна после входа в спинной мозг идут в составе дорсальных столбов и отдают коллатерали, оканчивающиеся на нейронах IV — VI пластин заднего рога. Некоторые волокна, делая в этой области восходящую петлю, входят в желатинозную субстанцию с вентральной стороны и разветвляются в радиальном направлении по всей ее глубине. Терминали этих волокон образуют синапсы на нейронах желатинозной субстанции и на дендритах глубжерасположенных клеток. В настоящее время очень мало известно о нейрохимической медиации в самом первом центральном синапсе: афферентное волокно-нейрон заднего рога спинного мозга. Предполагается несколько кандидатов, наиболее вероятными из которых считаются субстанция Р, холецистокинин, соматостатин, вазоактивный интестинальный полипептид.
Функциональная характеристика нейронов заднего рога, связанных с ноцицептивной афферентацией. Все нейроны дорсальной части заднего рога могут быть разделены на 3 группы. Первая состоит из клеток, активируемых исключительно ноцицептивными стимулами или раздражением «ноцицентивных» А-δ- и С-волокон. Вторая группа включает клетки, отвечающие на ноцицептивные и неноцицептнвные воздействия и на стимуляцию как высокопороговых, так и низкопороговых афферентов. Третья группа представлена нейронами желатинозной субстанции, которые, взаимодействуя с клетками первых двух групп, образуют систему модулирующего контроля афферентного входа в спинной мозг и формирования восходящего ноцицептивного потока импульсов.
Нейроны, активируемые ноцицептивными стимулами, так называемые специфические ноцицептивные нейроны локализованы в основном в I слое, где оканчиваются А-δ- и С-афференты. Они имеют ограниченные рецептивные поля и небольшую пространственную конвергенцию афферентных входов. Некоторые из них, как правило, не имеющие спонтанных разрядов, активируются только А-δ-волокнами, связанные с механорецепторами. Другие, спонтанно активные, отвечают на механическое, термическое ноцицептивное раздражение кожи и на электрическую стимуляцию А-δ- и С-волокон, причем возбуждающее действие С-волокон на эти клетки не зависит от соотношения активности в толстых и тонких немиелинизированных волокнах, так как ответ нейронов на тепловое ноцицептивное раздражение кожи очень мало изменялся при блоке проведения по А-волокнам.
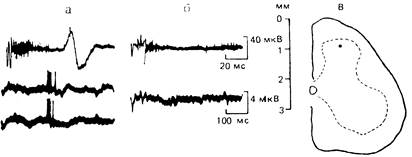
Рис. 2 - Ответ нейрона заднего рога спинного мозга на электрораздражение С-афферентов до, после поляризации афферентных волокон и локализация нейрона
По некоторым данным, более 25% нейронов желатинозной субстанции также реагировали исключительно на ноцицептивные стимулы. Предполагается, что такие клетки, активируемые С-волокнами, в функциональном отношении являются вставочными нейронами на пути передачи импульсов от первичных афферентов к нейронам I пластины. При одновременной регистрации активности пары клеток — одной в желатинозной субстанции, другой — в I пластине — было установлено, что латентный период первого потенциала действия в ответе нейрона желатинозной субстанции меньше, чем латентный период I спайка в ответной реакции нейрона I пластины и оба этих нейрона имели перекрывающиеся рецептивные поля. Эти различия были особенно характерны для нейронов наружной зоны желатинозной субстанции. Именно здесь располагаются клетки стеблевидной формы с аксонами, направляющимися в I пластину заднего рога. Кроме того, клетки желатинозной субстанции, активируемые С-афферентами, могут возбуждать нейроны не только I, но и V пластины. Различное региональное распределение клеток I пластины, которые получают ноцицептивный вход через А-δ-афференты, и клеток желатинозной субстанции, возбуждаемых С-волокиами, могут быть одной из причин возникновения «быстрой» и «медленной» боли, поскольку аксоны нейронов I пластины участвуют в формировании спиноталамической восходящей системы.
Предполагается, что «специфические» ноципептивные нейроны играют существенную роль в формировании восходящего ноцицептивного потока, который передает в высшие отделы мозга информацию об интенсивности и локализации ноцицептивных стимулов.
Однако ряд экспериментальных данных показывает, что роль клеток I слоя заднего рога, активируемых исключительно ноцицептивными афферентами, в общей картине процессов формирования восходящего болевого потока не должна излишне переоцениваться, как это было сразу после выявления этих «специфических» ноцицептивных нейронов Оказалось, что таких «специфических» нейронов не так уже много в I пластине и что они перемешаны с клетками, отвечающими на стимуляцию как высоко-, так и низкопороговых афферентов, а некоторые из них активируются и раздражением А β-афферентов и даже мы щечных волокон III и IV групп. Кроме того, эти клетки не активировались брадикинином, и их ответы на ноцицептивныс воздействия не коррелировали с поведенческим реагированием на боль. Выявлено, что аксоны нейронов I пластины проецируются не только в спиноталамический тракт, но и во все квадранты спинного мозга и даже в такой классически неболевой тракт, как дорсолатеральные канатики.
Нейроны, активируемые разномодальными афферентными входами, или «мультирецептивные», «конвергентные», «нейроны широкого афферентного диапазона», локализованные в основании заднего рога и глубже, играют основную роль в переработке ноцицептивной информации, в передаче ее в восходящем направлении и в реализации низкоинтегрированных простейших реакций на повреждающие воздействия. Строение, источники их активации, свойства этих клеток и проекции их аксонов хорошо исследованы.
Общепризнанно, что клетки основания заднего рога имеют достаточно широкое, хорошо контурируемое рецептивное поле, окруженное подпороговой зоной, имеющей незначительный афферентный вход на эти нейроны. В тех случаях, когда эффективность афферентного входа возрастает, рецептивное поле может увеличиваться. На этих нейронах конвергируют афферентные волокна различного спектра и модальности, поскольку они отвечают на прикосновение, давление, на механическую и термическую ноцицептивную стимуляцию рецептивного поля. Они моносинаптически активируются низкопороговымн А-волокнами и одновременно имеют полней наптический С-вход. Нейроны IV—V пластин отвечают высокочастотной «пачкой» импульсов на естественное неповреждающее раздражение кожи и длительной импульсной активностью при ноцицептинном воздействии. Частота и продолжительность ответов нейрона возрастает пропорционально интенсивности ноципептивных, особенно термических стимулов.
Нейроны V пластины реагируют с латентным периодом 15—30 мс длительной и высокочастотной импульсной активностью на внутриартериальное введение брадикинина, причем латентный период и продолжительность ответной реакции на брадикинин у приматов хорошо коррелируют с возникновением ощущения боли у человека.
Одновременное раздражение А- и С-афферентов проявляется двумя ответными реакциями клеток — ранней и поздней. Выключение проведения возбуждения по А-волокнам их поляризацией усиливает ответы клеток на раздражение С-волокон, и наоборот, предшествующее раздражение А-афферентов уменьшает их реагирование на ноцицептивное раздражение рецептивного поля или на активацию С-волокон. Нами было обнаружено, что ответная реакция нейронов V слоя на ноцицептивное механическое воздействие уменьшается, если оно наносится на фоне ритмической стимуляции низкопороговых афферентов. В последнее время получены весьма убедительные доказательства ведущей роли конвергентных нейронов в процессах кодирования интенсивности ноцицептивных термических стимулов. Предполагается, что изменение свойств конвергентных нейронов: рецептивного поля, спонтанной активности и ответов на разномодальные раздражения — у крыс с экспериментальным артритом лежат в основе гипералгезии и болевого синдрома у больных ревматоидным артритом.
На нейронах заднего рога конвергируют не только разномодальные кожные, но также мышечные и висцеральные афференты. Происходящее на этих нейронах взаимодействие соматовисцеральных афферентных потоков лежит в основе возникновения отраженной боли. Установлено, что нейроны, на которых осуществляется взаимодействие соматических и висцеральных афферентов, активируются через висцеральные входы при естественном раздражении висцеральных рецепторов, но только в том случае, когда интенсивность воздействия, например давление в желчных путях достигает ноцицептивного уровня.

Рис. 3 - Ответ нейрона IV слоя заднего рога спинного мозга на ноцицептивное раздражение кожи, наносимое до и после ритмической стимуляции пизкопороговых афферентов. 1 — фоновая импульсная активность: 2 — импульсная активность при ноцицептивном раздражении до стимуляции низкопороговых афферентов: 3 — то же после ритмического раздражения низкопороговых афферентов
Линия под осциллограммами — период ноцицсптпвного воздействия на кожу. Калибровка: времени — 1 с, амплитуды — 250 мкВ.
Естественно, что не все нейроны заднего рога имеют ноцицептивные афферентные входы. Во всех пластинах, особенно в IV, имеется достаточно большая популяция клеток с узким функциональным диапазоном, которые активируются легкими механическими воздействиями и не отвечают на повреждающие стимулы. В то же время клетки, отвечающие на ноцицепцию, выявлены в VI пластине и даже в вентральном роге. Эти нейроны, вероятно, участвуют в регуляции не только моторной, но и сенсорной деятельности спинного мозга, и в частности в регуляции боли, способствуя локализации ответа на боль. Поэтому цитоархитектонически выделяемые пластины в заднем роге спинного мозга скорее должны рассматриваться как зоны преимущественной концентрации нейронов с определенными свойствами, а не как строго функционально специализированные нейронные ансамбли.
Таким образом, можно считать, что конвергентные нейроны, локализованные в основании заднего рога, большинство из которых имеют восходящие проекции, принимают самое непосредственное участие в возникновении информации такого качества, которая высшими отделами головного мозга расценивается как болевая и которая запускает сложные механизмы ответного реагирования на боль. Однако в настоящее время есть все основания считать, что активность релейных нейронов, связанных с ноцицептивной афферентацией, их ответы на разномодальные стимулы, взаимодействие на них различных афферентных входов и, следовательно, формирование восходящего импульсного потока модулируется нейронами желатинозной субстанции.
Нейроны желатинозной субстанции, локализованные во II пластине, стали предметом активного исследования в последние 15—20 лет. Морфофункциональная организация этой области детально рассматривается в специальных обобщающих статьях и обзорах. Поэтому здесь мы коснемся лишь некоторых свойств нейронов желатинозной субстанции, необходимых для понимания их роли в регуляции ноцицепции.
Эти нейроны имеют рецептивные поля различных размеров, формы и локализации. У половины из них рецептивные поля по размерам и локализации соответствуют таковым у нейронов IV—V пластин и располагаются в пределах какой-либо одной области нижней конечности. Характерной особенностью 1/3 нейронов желатинозной субстанции является наличие у них очень маленьких рецептивных полей. У незначительного количества нейронов рецептивные поля занимали всю поверхность ипсилатеральной и иногда даже контралатеральной конечности. Именно на них конвергируют А-δ-, С-волокна, а на некоторых и А-р-афференты. Примечательно, что аксоны этих клеток не выходят за пределы спинного мозга.
Ориентация аксонов желатинозных нейронов весьма различна и во многом определяется локализацией сомы клетки. У нейронов, расположенных в дорсальной части желатинозной субстанции, аксоны направлены дорсально и проецируются в тракт Лиссауэра. Аксоны нейронов средней трети идут в продольном направлении внутри желатинозного вещества. По мере приближения к нижней границе желатинозной субстанции аксоны принимают вентральное направление и проникают, как правило, в глубжерасположенные слои заднего рога. О протяженности аксональных проекций имеются разноречивые и даже полярные представления. Согласно одним данным, аксоны нейронов желатинозной субстанции оканчиваются в пределах одного сегмента, в то время как по другим — они достигают даже таламуса. Однако аксоны большинства желатинозных нейронов не проецируются дальше одного сегмента и оканчиваются в пластинах I, IV, V или даже в пределах одной II пластины. У меньшей части нейронов аксоны, входящие в тракт Лиссауэра, прослеживаются на расстоянии 15 — 40 мм.
Отдельные клетки имеют контралатеральные аксональные проекции.
Основная масса нейронов имеет спонтанную активность в диапазоне от 1 до 40 имп/с. Она представлена разрядами постоянного ритма без высокочастотных «пачек» импульсов. Ни перерезка дорсолатеральных канатиков, ни полная спинализация существенно не изменяли фоновую активность желатинозных нейронов, что свидетельствует о ее независимости от нисходящей тонической регуляции и о большой роли афферентного сегментарного входа в ее формировании. Нейроны желатинозной субстанции реагируют на естественное раздражение их рецептивного поля. Большинство клеток отвечали как на неповреждающую механическую, так и на ноцицептивную стимуляцию. По данным Р. Cervero и A. Iggo, разномодальная активация рецептивного поля сопровождается исключительно торможением активности нейронов, и в зависимости от характера воздействия они могут быть разделены на 3 группы. В первую группу входят клетки, активность которых тормозилась при низкопороговой механической стимуляции рецептивного поля. Вторую группу составляют нейроны, торможение активности которых возникало при раздражении рецептивного поля как повреждающими, так и неповреждающими стимулами. К третьей группе были отнесены клетки, разряды которых угнетались только при ноцицептивном раздражении рецептивного поля. Эти 3 группы клеток в функциональном отношении, т.е. по характеру ответов на разномодальную стимуляцию рецептивного поля, являются антиподами глубжерасположенных нейронов.
По общепринятому мнению, клетки желатинозной субстанции отличаются от других нейронов заднего рога, в частности V пластины, характером ответов на раздражение С-волокон и взаимодействием А- и С-афферентных входов. Так, нейронам желатинозной субстанции не свойствен феномен «взвинчивания». Они не изменяют, как нейроны V пластины, ответы на раздражение С-волокон при выключении А-афферентов, а предшествующая стимуляция их не уменьшает ответные реакции желатинозных нейронов, возникающие при внутриартериальном введении брадикинина. Наряду с хорошо изученным быстрым реагированием нейронов желатинозной субстанции на разномодальные афферентные воздействия, выявлены более медленно и длительно протекающие изменения их активности. Эти изменения, вероятно, обусловлены действием различных биологически активных соединений, осуществляющих нейромодуляторную функцию, таких как энкефалпны, соматостатин, ангиотензин, холецистокинин, гликокортикоиды, субстанция Р, ГАМ.К, норадреналин, серотонин, локализующихся в дорсальной части заднего спинного мозга. Однако функциональная роль медленных, длительных изменений активности нейронов желатинозной субстанции и их связь с регуляцией ноцицептивного афферентного потока пока не выяснены и требуют новой концептуальной гипотезы.
Функциональная роль желатинозной субстанции до настоящего времени весьма спорна. Практически все исследователи придают ей важное значение как нейрональной системе, регулирующей сенсорный вход и болевую чувствительность на сегментарном уровне. Однако каждый имеет свое, порой весьма противоречащее имеющимся, представление о механизмах этой регуляции. В настоящее время снова получила подтверждение и развитие точка зрения, высказанная более 30 лет назад, о том, что желатинозная субстанция функционирует как релейная инстанция, передающая ноцицептивную информацию от первичных афферентов к нейронам спиноталамического тракта. Именно в области желатинозной субстанции оканчивается основная масса ноцицептивных А-δ- и С-афферентов, которые непосредственно активируют нейроны желатинозной субстанции.
Согласно другой, более общепринятой концепции клетки желатинозной субстанции не просто передают, а модулируют, эффективность синаптического воздействия афферентных входов на нейроны дорсального рога спинного мозга. Традиционно считается, что эта модуляция реализуется тормозными механизмами и что большая часть клеток желатинозной субстанции функционирует как тормозные интернейроны. Примечательно, что налоксон при внутривенном введении, возбуждая нейроны V пластины, одновременно угнетает активность нейронов желатинозной субстанции.
Модуляция афферентного сегментарного входа клетками желатинозной субстанции может осуществляться как пост-, так и пресинаптическими механизмами. В настоящее время известно немало достаточно хорошо проверенных фактов, свидетельствующих о важной роли желатинозной субстанции в генерации деполяризации первичных афферентов. Желатинозной субстанции отводится ключевая роль в одной из самых распространенных и, пожалуй, самых дискуссионных теорий боли — теории «входных ворот» P. Мелзака и Р. Уолла. Эта теория в первом ее постулате объясняет формирование ноцицептивного потока на сегментарном уровне на основе центрального взаимодействия быстропроводящей миелиновой и медленнопроводящей безмиелиновой систем на первых релейных нейронах спинного мозга. Афферентная импульсация, возникающая в миелиновых волокнах при неповреждающих раздражениях, одновременно с активацией релейных нейронов через коллатерали возбуждает нейроны желатинозной субстанции. Они, в свою очередь, деполяризуют первичные миелиновые афференты и тем самым пресинаптически ограничивают активирующее их влияние на Т-клетки. При ноцицептивном воздействии активируются тонкие миелиновые и безмиелиновые волокна, которые оказывают тормозное влияние на нейроны желатинозной субстанции, в результате чего уменьшается деполяризация миелиновых волокон, т.е. «открывается» афферентный вход и возрастает эффективность синаптического воздействия на Т-нейроны. Т-нейроны при достижении критического уровня, определяемого временной и пространственной суммацией «ноцицептивных» и неноцицептивных сигналов, формируют восходящий поток импульсов нового качества, который воспринимается вышерасположенными структурами мозга.
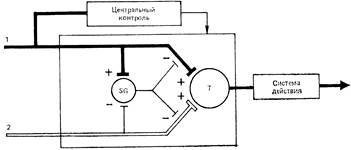
Рис. 4 - Схема системы контроля афферентного входа
Положение теории Р. Мелзака и П. Уолла о том, что процессы пресинаптического торможения и облегчения являются ведущими в реализации модулирующего влияния желатинозной субстанции, длительное время являлись предметом активного обсуждения. Эта теория, как и любая новая теория, постоянно совершенствуется и дополняется. В современных ее модификациях, и в том числе самих авторов, больший акцент сделан на роль постсинаптических и нейрохимических процессов в регуляции передачи разномодальных импульсов на релейные нейроны спинного мозга.

Рис. 5 - Схема лемнисковой и экстралемнисковой восходящих афферентных систем
Таким образом, можно считать, что формирование восходящего «ноцицептивного» потока в спинном мозге происходит двумя способами — на основании взаимодействия разномодальных афферентных входов на релейных нейронах и в результате активации «специфических болевых» нейронов, имеющих восходящие проекции.
Восходящие пути болевой чувствительности. Существуют 2 основные «классические» — лемнисковые и экстралемнисковые — восходящие афферентные системы. В пределах спинного мозга одна из них располагается в дорсальной и дорсолатеральной зоне белого вещества, другая — в его вентролатеральной части. Известно, что в ЦНС не существует специализированных путей болевой чувствительности и что интеграция боли осуществляется на различных уровнях ЦНС на основе сложного взаимодействия лемнисковых и экстралемнисковых проекций. Вместе с тем не вызывает сомнений тот факт, что значительно большую роль в передаче восходящей ноцицептивной информации играют вентролатеральные проекции, морфофункциональная организация которых детально исследована и обобщена в ряде монографий и обзоров. Об этом свидетельствуют и клинические наблюдения, показывающие, что после антеролатеральной хордотомии или деструкции этой системы при некоторых заболеваниях возникает длительная и выраженная аналгезия. С другой стороны, ощущение боли и ее локализация сохраняются при перерезке всех, кроме вентролатеральных, путей, а стимуляция аксонов в антеролатеральном квадранте после хордотомии вызывает интенсивную боль.
Источниками вентролатеральных проекций являются нейроны I—V и даже VI—VIII пластин спинного мозга, аксоны которых имеют диаметр от 1 до 11 мкм и скорость проведения 7—74 м/с. Вентролатеральные восходящие пути в пределах спинного мозга имеют определенную соматотопическую организацию — волокна, исходящие из более каудальных сегментов, располагаются более латерально, а из ростральных — медиально и вентрально. Конечные проекции первоначально выявляются в п