Раздел 1. Биологическая эволюция
Концепции происхождения жизни
 Существует несколько гипотез, объясняющих возникновение жизни:
Существует несколько гипотез, объясняющих возникновение жизни:
Согласно гипотезе стационарного состояния виды неизменны, Земля существовала вечно и всегда была способна поддерживать жизнь. В пользу этой теории свидетельствуют разногласия в оценке возраста Земли и разрывы в палеонтологической летописи, против нее – все свидетельства глобальной эволюции. Гипотеза панспермии утверждает, что жизнь могла попасть на Землю с других планет в разных частях Вселенной. В пользу этой теории говорит наличие «предшественников живого» – цианогенов, синильной кислоты и органических соединений или даже следов примитивных форм жизни на метеоритных остатках. Недостаток теории в том, что она не объясняет происхождение жизни на других планетах. Согласно креационизму, жизнь возникла в результате вмешательства сверхъестественного существа[1]. Однако знание, полученное через божественное откровение и веру, является субъективным, не подтверждается экспериментально, не может быть объектом научного исследования. Сверхъестественное объяснение происхождения жизни относится к ответу, скорее, на вопрос «почему», а не «каким образом». Как и первые две теории, креационизм не объясняет механизмов биогенеза. Гипотеза самозарождения утверждает наличие животворного активного начала в неживых объектах. В связи с этой гипотезой обычно вспоминают «удачный» трехнедельный эксперимент нидерландского каббалиста и алхимика Жана Батиста ван Гельмонта (1577–1644) по созданию мышей из грязной ветоши и пшеницы, помещенных в корзине в темный шкаф[2], и «неудачный» эксперимент флорентийского врача Франческо Реди (1626–1697) по выведению мух из мяса, помещенного в сосуды с затянутым кисеей горлышком. Два века понадобилось Ладзаро Спалланцани (1765) и Луи Пастеру (1860), чтобы распространить постулат «живое только из живого» на микроорганизмы, которые не пережили их опытов с кипячением и пастеризацией.
Согласно гипотезе биохимической эволюции Опарина – Холдейна (1923) развитие жизни (биосферы) является следствием длительной химической эволюции – абиогенеза, и неразрывно связано с формированием и изменением других оболочек Земли – литосферы, гидросферы, атмосферы. Данная гипотеза считается самой обоснованной.
Абиогенез
ДОГЕОЛОГИЧЕСКИЙ ПЕРИОД. По современным радиометрическим оценкам, возраст Земли составляет около 5000×106 лет. Полагают, что в то время планета была не такой плотной, ее гравитационное поле было слабее. Постепенно произошла гравитационная стратификация, сформировались геосферы. По мере дифференциации и дегазации геосфер сформировались гидросфера и атмосфера. Легкие газы (водород, гелий, азот, кислород, аргон) при слабой гравитации улетучивались, но их более тяжелые соединения (вода, аммиак, двуокись углерода, метан) удерживались гравитацией. Древняя атмосфера (как и атмосферы других планет) была бескислородной, восстановительной, на что указывают "кислородофобные" минералы (обломочный пирит) и двухвалентное восстановленное железо (FeО), в отложениях того возраста (и позже, до 2,2 млрд. лет назад)[3].
Самые тяжелые элементы погружались к центру Земли и примерно 3,6 млрд лет назад сформировали твердое внутреннее ядро (80% Fe + 20% Ni), окруженное внешним ядром (88%Fe + 12% S). Над ним расположен слой мантии (43% SiO2 + MgO2 + Fe2O3 + CaO), а над ней – земная кора (50% SiO2 + 15% Al2O3), Земная кора состоит из магматических, осадочных и метаморфических горных пород, относительный или абсолютный возраст которых можно установить по наличию руководящих ископаемых, составу, глубине залегания[4]. Самыми древними являются наиболее глубокие породы – 8-километровый слой базальта в океанической коре или 35-70-километровый слой базальта и гранита в континентальной.

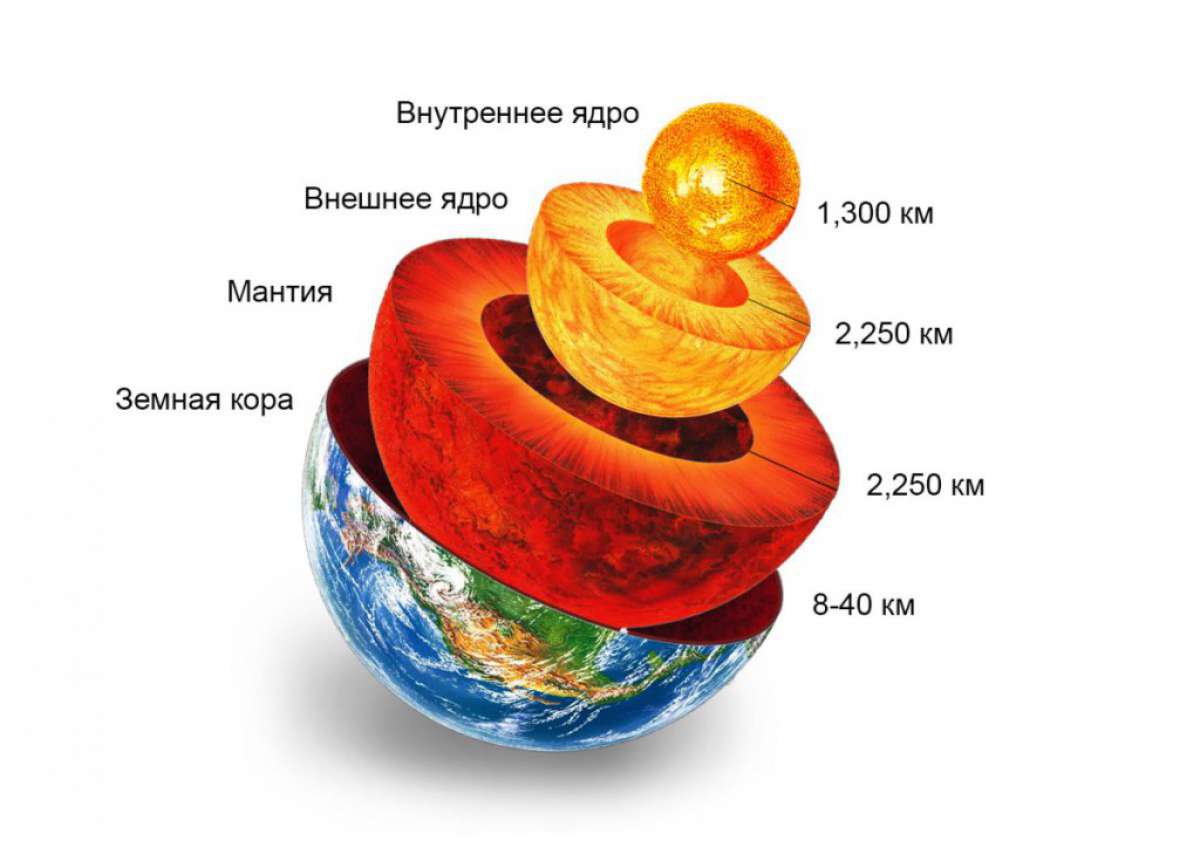
Вместе с верхней мантией кора образует литосферу, состоящую из малых, средних и 8 больших тектонических плит. В ядре и мантии постоянно происходит конвекция (перемешивание) остывших верхних и горячих внутренних слоев, и конвективные потоки сдвигают литосферные плиты. В зоне спреддинга («раздвигания» плит) образуется рифт – новая океаническая кора (Срединно-Атлантический хребет). В зоне субдукции («подминания» плит) образуются горы и острова (Анды, Кордильеры, Камчатка, Курилы, Гималаи). В результате этого дрейфа (от нескольких мм до 8 см/год) с периодичностью 600-800 миллионов лет (цикл Вильсона) образуются суперконтиненты. Последний суперконтинент – Пангея, сформировался приблизительно 250 млн лет назад на границе палеозоя и мезозоя и просуществовал около 50 млн лет. На данный момент Атлантика расширяется, а Тихий океан закрывается, Индостан сближается с Евразийской плитой. Предполагают, что через 20 млн лет сольются Африка и Европа, через 60 млн лет к ним примкнет Австралия, через 130 млн лет – Антарктида. Через 200 млн лет новый суперконтинент закончит свое формирование.

Абиогенез
Историю Земли разделяют на эоны, эры и периоды.


КАТАРХЕЙ. 4,5 – 4 млрд лет назад происходит стратификация планеты и формирование спутника (предположительно, в результате столкновения с Землей какого-то крупного небесного тела). Ранее считалось, что поверхность Земли была в то время раскаленной и непригодной для жизни. Сейчас предполагают, что несмотря на высокую вулканическую активность литосфера и гидросфера были уже сформированы, что согласуется с возрастом циркона (ZrSiO4), обнаруженного в Канаде и Австралии. Интересно, что включения графита и алмазов в катархейских апатитах и цирконе содержат неожиданно много легких изотопов углерода 12С, что является косвенным свидетельством наличия жизни[5]. Приблизительно 4,1 – 3,8 млрд лет назад Луна и Земля подверглись «поздней тяжелой бомбардировке» метеоритами, практически вся катархейская кора переплавилась. Уничтожили эти метеориты первое поколение живых существ, принесли его с собой или пустыми упали на тогда еще пустую планету – остается загадкой.
АРХЕЙ. 4 – 2,5 млрд лет назад. 3,8 млрд лет назад заканчивается метеоритная бомбардировка, снижается вулканическая активность, завершается формирование твердого ядра и континентальной коры; появляются первые осадочные породы (запад Гренландии), кол-во восстановленного железа уменьшается. Начинается «читаемая» геологическая летопись, в которой признаки жизни пока ограничиваются облегченным изотопным составом углерода3. Согласно гипотезе А.И. Опарина, архейская атмосфера и гидросфера оказалась благоприятной для синтеза простых органических соединений (метана, аммиака) из неорганики. Поскольку реакции синтеза являются эндергоническими, они невозможны без внешних источников энергии; предполагается, что такими источниками послужили электромагнитная энергия солнечного света, тепловая энергия вулканических извержений, химическая энергия окислительных реакций. Катализатором служили твердые частицы, содержащие железо и никель. В экспериментах с использованием установки Стэнли Миллера (1953), воспроизводящих условия древнего океана, удалось наблюдать самосборку ряда аминокислот, аденина, простых сахаров (рибозы), а позже – шестимономерных нуклеиновых кислот.
Живые растворы. Ранний абиогенез протекал, вероятнее всего, в специфических средах, таких как поверхности глинистых минералов мелководья, микрополости кристаллов пирита, ячейки породы подводных вулканических выходов.[6] Минералы обеспечивали: 1. Накопление, концентрацию реагентов. Например, цеолиты, обладающие своеобразной кристаллической решеткой, способны пропускать мелкие и задерживать крупные ионы и молекулы. Слоистые кристаллические решетки глинистых минералов накапливают, концентрируют в межслоевых пространствах короткие молекулы аминокислот. 2. Катализ. Например, магнетит вызывает рекомбинацию атомов азота и водорода с образованием аммиака, из которого легче извлекается азот. Пирит содержит распространенные кофакторы − железо и серу. 3. Пространственную организацию реагентов и катализаторов. Например, кальцит обладает избирательностью разных граней кристалла в отношении "левых" и "правых" молекул аминокислот. 4. Источник энергии. Например, пирит может быть донором свободных электронов.
Первостепенная роль в процессах происхождения жизни приписывается: 1. Переносчикам протонов, обеспечивающим обмен энергией. Во всех живых клетках основным источником энергии служит АТФ – аденозинтрифосфорная кислота. Более специфичны ГТФ, ЦТФ, УТФ, НАД+. Предполагается, что так было не всегда, и у архаичных видов роль последних в обмене выше. 2. Катализаторам, ускоряющим определенные реакции. Не исключено, что на ранних этапах становления жизни в качестве катализатора и источника электронов служили ионы металлов, в том числе тяжелых. В современных клетках они выполняют роль коферментов.[7] Возможно, первые ферменты обладали большим разнообразием сравнительно с современными. 3. Информационным молекулам, обеспечивающим наследственность. Самые первые из них – рибозимы – состоящие из РНК РНК-зависимые РНК-полимеразы, т.е. РНК, способные катализировать синтез собственных копий. В искусственных условиях получены прообразы РНК-организма – самовоспроизводящиеся колонии рибозимов, катализирующие синтез нуклеотидов, присоединяющие аминокислоты к РНК, синтезирующие белки и НАД. Как и у белков, их активность повышается при присоединении металлов.[8] Со временем между белками и НК формировались кросс-каталитические отношения, в которых белки выполняли роль катализаторов синтеза НК, а НК – матриц для синтеза белков.[9]
Абиогенное формирование органических молекул, их отбор и усложнение происходили на протяжении почти 1 миллиарда лет. Все это время стабилизировались и циклически повторялись реакции с частичным возобновлением расходуемого субстрата, кросс-каталитические и автокаталитические реакции (образование сахаров из формальдегида, самосборка рибозимов и др.). Таким образом в результате абиогенеза формируется «мир РНК», «первичный бульон», «живые растворы» – многокомпонентные химические системы, поддерживающие постоянство внутренней среды, только без внутренней среды J. Живая материя приобретает свое первое свойство – цикличность.
 Коацерватные капли («сгустки»). Вопрос пространственного ограничения гиперциклов до сих пор остается открытым. Возможно: 1) результатом абиогенеза были белково-нуклеотидные сгустки с общей гидратной оболочкой, а уже на границе водораздела сформировались билипидные мембраны[10]; 2) первые коацерваты образовались самопроизвольно из абиогенных липидов, а впоследствии вступили в симбиоз с колониями РНК и рибозимов, среди которых были и катализаторы синтеза липидов и белков[11].
Коацерватные капли («сгустки»). Вопрос пространственного ограничения гиперциклов до сих пор остается открытым. Возможно: 1) результатом абиогенеза были белково-нуклеотидные сгустки с общей гидратной оболочкой, а уже на границе водораздела сформировались билипидные мембраны[10]; 2) первые коацерваты образовались самопроизвольно из абиогенных липидов, а впоследствии вступили в симбиоз с колониями РНК и рибозимов, среди которых были и катализаторы синтеза липидов и белков[11].
Наличие мембран позволяет: 1. Избирательно накапливать вещества внутри пузырьков. 2. Создавать локальные градиенты концентрации за счет каталитического превращения веществ, а возможно, и за счет активного транспорта. 3. Действовать групповому отбору. Основная эволюционная тенденция того времени заключалась в ускорении реакций, и преимущество получали рибозимы с высокой скоростью саморепликации, т.е. короткие (монстр Шпигельмана). Однако они обладают слабой каталитической активностью, и размножаясь, губят всю колонию (как если бы в колонии пчел остались одни трутни). С момента коацервации индивидуальный отбор, направленный на отдельные молекулы, дополняется групповым, направленным на сообщества репликаторов, заключенные в пузырьки (подробнее) (и еще) (и еще). «Выживают» сообщества, где каждый полезен не только для себя (см. «Парадокс Симпсона», «Эволюционно стабильная стратегия», или «Равновесие Нэша», или «Дилемма заключенного», или «Игры разума» J).
Таким образом, за «живыми растворами» следуют коацерватные капли – липидные пузырьки, содержащие белки и НК, способные поддерживать постоянство внутренней среды, а в определенных условиях расти и делиться (но не размножаться). Живая материя приобретаетдискретность.
| Процесс передачи информации | Синтези-руется | Матри-ца |
| 1. Репликация | ДНК | ДНК |
| 2. Транскрипция | РНК | ДНК |
| 3. Репликация РНК | РНК | РНК |
| 4. Обратная транскрипция | ДНК | РНК |


 Постепенно происходит передача функций носителя информации от РНК к ДНК, не способной к «самообслуживанию», но более стабильной и надежной. Существует несколько гипотез о причинах такой «смены власти». 1. РНК-жизнь существовала в оптимуме температур около 20º (или потому, что возникла до «поздней тяжелой бомбардировки», или потому, что возникла на полюсах). При разогреве планеты (или при распространении к экватору) становится выгоднее использовать ДНК, для которой оптимум составляет около 60º (подробнее). 2. ДНК была покоящейся фазой в жизненном цикле рибозимов. 3. ДНК более устойчива к атакам паразитов.
Постепенно происходит передача функций носителя информации от РНК к ДНК, не способной к «самообслуживанию», но более стабильной и надежной. Существует несколько гипотез о причинах такой «смены власти». 1. РНК-жизнь существовала в оптимуме температур около 20º (или потому, что возникла до «поздней тяжелой бомбардировки», или потому, что возникла на полюсах). При разогреве планеты (или при распространении к экватору) становится выгоднее использовать ДНК, для которой оптимум составляет около 60º (подробнее). 2. ДНК была покоящейся фазой в жизненном цикле рибозимов. 3. ДНК более устойчива к атакам паразитов.

| ДДП, 17,8% |
| Alu, 11,2% |
| SVA, 0,2% |
| Прочие, 6,3% |
| ДКП, 8,8% |
| ДНК-транс-позоны, 3% |
| Неперемеща- емые части генома, в т.ч. протеинко-дирующие последова-тельности 52,7% |
1. Репликация – точное копирование НК по принципу комплементарности. Используется про- и эукариотами для удвоения ДНК при делении, дву- и однонитевыми ДНК-вирусами (вирус гепатита В, герпеса, парвовирус) [подробнее].
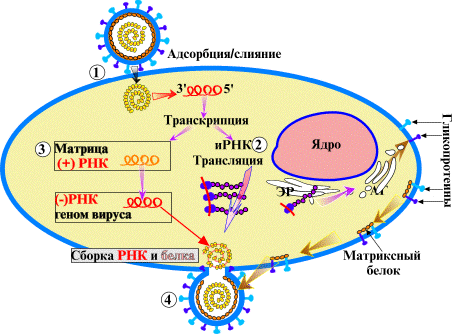
2. Транскрипция – синтез РНК на основе ДНК по принципу комплементарности. Так синтезируется мРНК у клеток и вирусов, РНК-прегеном у вируса гепатита В (тем и уникален).
3. Репликация РНК – точное копирование РНК, предположительно самый древний механизм воспроизводства, характерный для колоний рибозимов. Сейчас используется одно- и двунитевыми РНК-вирусами для синтеза иРНК и собственного генома, а также «монстром Шпигельмана» для саморепликации J. У про- и эукариот неизвестен.
4. Обратная транскрипция – синтез ДНК на основе РНК. Используется ретровирусами (например, ВИЧ, Т-клеточной лимформы) и катализируется обратной транскриптазой (ревертазой), которую вирус приносит с собой[12]. Кроме вирусов, такой способ самовоспроизведения используют некоторые разновидности мобильных генетических элементов. МГЭ – «транспозоны», «псевдогены», «гентические паразиты», – фрагмены ДНК, которые могут перемещаться и размножаться внутри генома. К ним относятся транспозоны, случайно интегрированные в геном бактериофаги, интроны и др. У человека МГЭ составляют до 45 % всей ДНК, у плодовой мухи 15-20 % у кукурузы 85 %. ДНК-транспозоны для перемещения применяют метод «вырезать и вставить», т.е. просто перемещаются, а ретротранспозоны – метод «копировать и вставить», т.е. еще и размножаются, используя прямую и обратную транскрипцию. Они повышают траты энергии на репликацию, синтез ревертазы и др. белков, повышают риск мутаций (в т.ч. и из-за низкой «аккуратности» ревертазы); поэтому их экспрессия жестко контролируется (особенно во время эмбриогенеза и гаметогенеза).
 5. Трансляция – синтез белка из аминокислот на основе мРНК, включает 3 этапа: 1. Инициация. тРНК, несущая метионин, узнает стартовый кодон (AUG) и присоединяется к нему; вокруг них из большой и малой субъединиц собирается рибосома. 2. Элонгация. К следующему кодону присоединяется соответствующая ему тРНК, а ее аминокислота – к предыдущей; рибосома перемещается вдоль мРНК и цикл многократно повторяется. 3. Терминация. Рибосома доходит до стоп-кодона (в данном случае UAG), и новосинтезированный белок отсоединяется от рибосомы (подробнее).
5. Трансляция – синтез белка из аминокислот на основе мРНК, включает 3 этапа: 1. Инициация. тРНК, несущая метионин, узнает стартовый кодон (AUG) и присоединяется к нему; вокруг них из большой и малой субъединиц собирается рибосома. 2. Элонгация. К следующему кодону присоединяется соответствующая ему тРНК, а ее аминокислота – к предыдущей; рибосома перемещается вдоль мРНК и цикл многократно повторяется. 3. Терминация. Рибосома доходит до стоп-кодона (в данном случае UAG), и новосинтезированный белок отсоединяется от рибосомы (подробнее).
Механизмы воспроизводства НК, а особенно их вариации у вирусов и МГЭ, позволяют предположить, что ранняя жизнь была в некоторых смыслах еще разнообразнее и удивительнее, чем нынешняя. Постепенно происходит унификация, и ДНК становится единственным способом закреплять информацию о произошедших изменениях (у всех, кроме вирусов). Иными словами, ДНК становится универсальной библиотекой генетической памяти. А РНК – библиотекарями J. Живая материя приобретает память.
| Наука | Определение жизни | Свойства жизни |
| Химия | фазообособленная форма существования автокатализаторов, | способных к мутациям и эволюции за счет естественного отбора. |
| Физика | открытые, саморегулирующиеся и самовоспроизводящиеся биополимерные системы, | с вектором развития, направленным на уменьшение собственной энтропии. |
| Экология | процессы сопряженной циркуляции биоэлементов в водной среде за счет энергии солнечного излучения, | способные к поступательному усложнению. |
| Кибернетика | высокоустойчивое состояние вещества, | использующее для автостабилизации информацию, кодируемую состояниями отдельных молекул. |
| Определение NASA | самоподдерживающаяся химическая система, | способная к дарвиновской эволюции. |
Обобщая все сказанное, жизнь – это самовоспроизведение с закреплением изменений, жизнь эмерджентна, непрерывна и необратима.
Биогенез
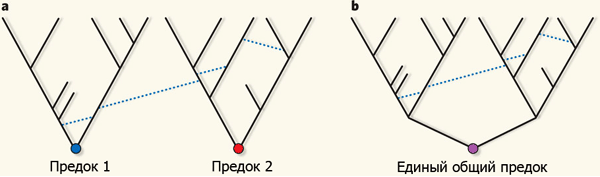
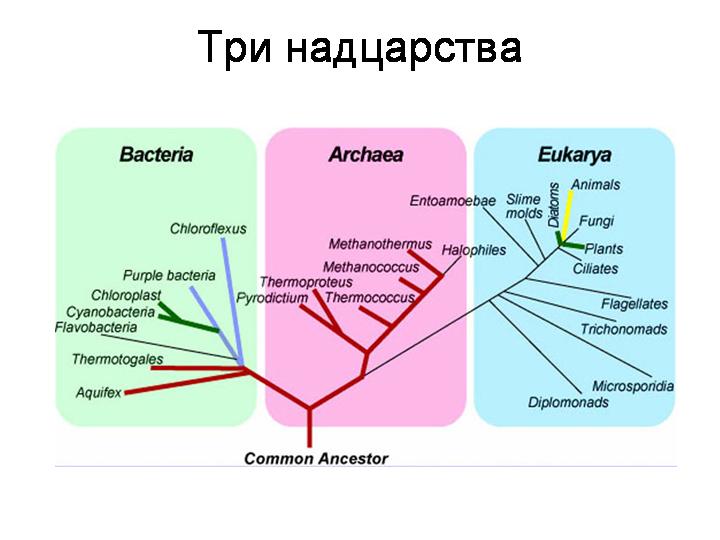

LUCA (last universal common ancestor) – последний общий предок. Анализ количества мутаций в генах, кодирующих рРНК, РНК-полимеразу и ряд других белков у представителей разных таксонов (молекулярные часы), позволил оценить относительную давность большинства ароморфозов и построить общее филогенетическое дерево, а также показал основные характеристики, общие для всех живых (и только живых) организмов. Объединив эти свойства, получили «образ» гипотетического универсального предка (и еще) (и еще):
· Он окружен билипидной мембраной и использует активный транспорт [13]. В качестве источника энергии использует глюкозу (D-изомер) или, по другим данным, Н2. В качестве переносчика энергии использует АТФ, которая синтезируется АТФ-синтазой за счет трансмембранного градиента протонов. Он анаэроб и термофил.
· Он кодирует информацию в ДНК. Генетическая информация содержится в двунитевой ДНК, состоящей из четырёх нуклеотидов (аденин, гуанин, тимин, цитозин). Репарация ДНК обеспечивается группой ферментов (топоизомераза, ДНК-лигаза и др.). РНК производится ДНК-зависимой РНК-полимеразой, вместо тимидина использующей уридин.
· Он синтезирует белки. НК обеспечивают синтез белков, а белки − синтез липидов и углеводов. Белки собираются из свободных аминокислот РНК (мРНК, рРНК, тРНК) и белками. Используются только L-изомеры аминокислот, которые синтезируются из D-изомеров глюкозы.
· Он имеет современные рибосомы из РНК и белков. Поступательное усложнение их структуры (добавление второй субъединицы, «ручек» для захвата тРНК) скорее всего, связано с увеличением точности транскрипции (подробнее).
· Он использует современный генетический код. Код избыточен – только 20 аминокислот кодируются 64 триплетными кодонами. У эукариот и прокариот код одинаковый, у архей и митохондрий немного отличается.
· Он размножается (пока только бесполым путем). При этом происходит вначале репродуцирование всего содержимого, затем деление.
Образ Луки, созданный филогенистами, остается неполным по нескольким причинам. 1. У всех живых организмов общими оказалось меньше генов, чем необходимо для жизни прокариота (минимум 400−500). Неизвестно, кому наш предок передал уникальные гены, и передал ли он их хотя бы кому-то. 2. Не обнаружилось общих генов, обеспечивающих репликацию ДНК и биохимию клеточной мембраны. Возможно, у Луки и не было ни ДНК, ни мембраны (подробнее).
Существует 2 возможных объяснения этих противоречий: 1. LUCA был не клеткой, а клетками, способными к горизонтальному переносу генов. В этом случае все особенности отдельных доменов могли быть получены от других организмов, существовавших одновременно с Лукой, но не выдержавших конкуренции и не прошедших через «бутылочное горлышко» эволюции. В пользу этого допущения свидетельствует и тот факт, что устойчивое существование возможно только при условии разнообразия. Однотипные живые существа очень быстро израсходуют все ресурсы или отравят себя продуктами собственной жизнедеятельности. 2. LUCA был не клеткой, а РНК-сообществом (Last Universal Common Ancestor State или LUCAS J). В этом сценарии ДНК-геном и мембрана возникли уже после разделения главных ветвей жизни. Следует учитывать, что оба объяснения не исключают друг друга и непротиворечивы только с научной точки зрения[14].
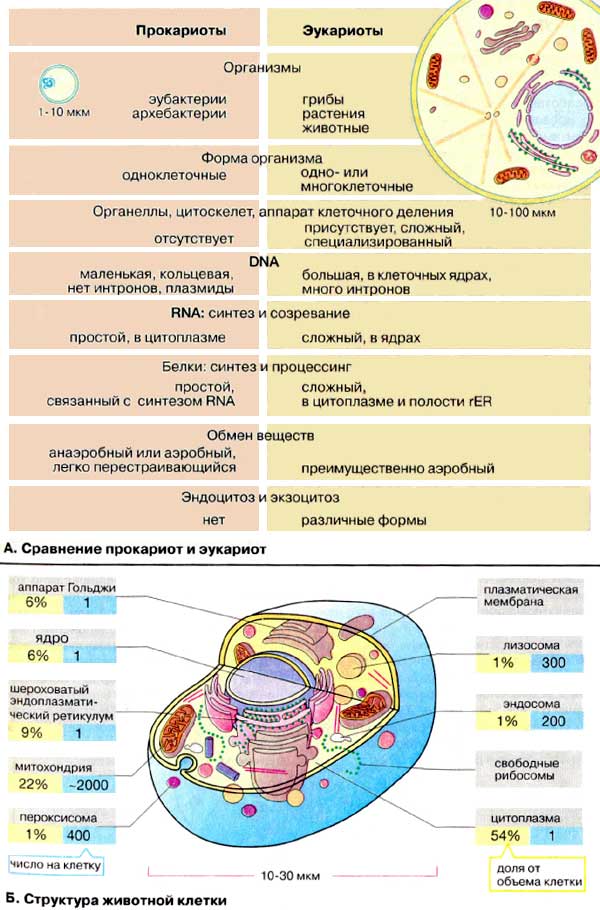 ПРОКАРИОТЫ (еще) (доя́дерные от греч. πρό «перед» и κάρυον «ядро»):
ПРОКАРИОТЫ (еще) (доя́дерные от греч. πρό «перед» и κάρυον «ядро»):
· Маленькие (1–10 мкм), не имеют цитоскелета и мембранных органоидов (за исключением плоских цистерн у фотосинтетиков, например, у цианобактерий), зато имеют жгутики и газовые вакуоли, клеточную стенку, иногда – капсулу. Двухцепочечная ДНК не содержит интронов, упакована без участия гистонов в плазмиды и нуклеоид – кольцевую или линейную хромосому. РНК синтезируются в цитоплазме, сложный аппарат репликации и транскрипции отсутствует.
· Размножаются делением, фрагментацией, почкованием; активно используют непосредственный (конъюгация, трансформация) или вирус-опосредованный (трансдукция) горизонтальный перенос генов, поэтому понятие вида к ним неприменимо.
· Питаются только посредством пиноцитоза. Неблагоприятные условия переживают в виде спор, многие могут перестраивать обмен веществ. Никогда не бывают многоклеточными. Никогда не бывают паразитами.
Раньше самыми древними прокариотами считались археи. Но интересно, что для них характерны некоторые прогрессивные признаки эукариот. Так, ДНК связана с гистонами, есть интроны и процессинг РНК. Похожи на эукариотические АТФаза, РНК-полимераза, ДНК-полимераза, промоторы и другие компоненты транскрипционного комплекса; процесс репликации и репарации ДНК. Еще одно доказательство близости архей и эукариот – это вид архей, названный Локи (обнаружен впервые в 2015 в гидротермальных полях Северной Атлантики – «замке Локи»), и его родственники – Тор, Один и Хеймдаль, объединенные в надтип Асгард J. Помимо вышеназванных «эукариотических» признаков, для асгардархей характерны многие компоненты цитоскелета, механизмы везикулярного транспорта, деградации белков. Значит, эволюционная модель «Лука ⇒ археи ⇒ бактерии ⇒ эукариоты» несостоятельна, ее следовало бы заменить на «Лука ⇒ бактерии ⇒ археи ⇒ эукариоты». Но в рамки второй модели не вписываются свойства, характерные только для архей – особенности клеточной стенки, наличие в мембране липидов с высокомолекулярными спиртами вместо жирных кислот. На данный момент рассматриваются две модели: «Лука ⇒ бактерии + археи + эукариоты» и «Лука ⇒ бактерии + археи ⇒ эукариоты».
Последовательность появления различных экологических групп прокариот остается невыясненной. Известно, что все бактерии способны фиксировать свободный СО2 (т.е. питаться автотрофно), однако большинство использует более экономный, гетеротрофный тип питания (получают биогеную органику). Автотрофное питание в широком смысле этого слова (как процесс синтеза органики) является более древним, однако неясно, кто именно совершил переход к гетеротрофии – «живые растворы», коацерваты или уже живые прокариоты.
| Источник органики | Источник энергии | |
| УФ | ОВР | |
| Синтезирует | Фотоавтотроф: -лито- (растения, лишайники, цианобактерии, зеленые и пурпурные серобактерии); -органо- (некоторые пурпурные бактерии, эвгленовые). | Хемоавтотроф: -лито- (большинство бактерий); -органо- (некоторые метаногены, денитрификаторы). |
| Поглощает | Фотогетеротроф: -лито- (некоторые цианобактерии, пурпурные, зеленые серобактерии); -органо- (галобактерии, пурпурные несерные бактерии) | Хемогетеротроф: -лито- (некоторые сульфат-редукторы); -органо- (животные, грибы, бактерии-паразиты, симбионты, паразиты). |
Анаэробные хемотрофы. Если связывать накопление О2 в атмосфере именно с деятельностью биоты, необходимо признать, что первые ее представители были облигатными анаэробами, т.е. могли жить только в бескислородной среде. Скорее всего, это были гипертермофильные хемотрофы, а именно, гидрогенотрофы, использующие в качестве восстановителя эндогенный Н2, образованный при реакции Н2О с перегретыми горными породами (например, сульфат-редукторы и метаногены). Разделение хемотрофов на авто- и гетеротрофов приводит к усложнению и удлинению пищевых цепей. Появляется новый тип взаимодействия – экологический.
Анаэробные фототрофы. По мере понижения уровня моря при формировании первого континента, обеднения минерального состава рифтовой зоны, повышения конкуренции за органику, прокариоты начинают использовать для синтеза органики солнечную энергию (например, пурпурные бактерии). Произошла тихая подводная революция, открытие нового источника энергии – солнечного света; фотосинтетики включили Солнце в свою окружающую среду, тем самым укрепляя базу для развития биосферы.
Аэробные фототрофы. В промежутке от 3,5 до 2,5 млрд появляются цианобактерии, они же – сине-зеленые водоросли[15]. Крупные, морфологически сложные прокариоты, содержат тилакоиды, обладают циркадным ритмом, «изобрели» симбиоз и оксигенный фотосинтез (фотохимическое расщепление воды). Обладая способностью к фотосинтезу, они играют важную роль в формировании первых сложно организованных экосистем – многослойных бактериальных матов (3,55 млрд лет назад). Верхние светопроницаемые слои в них заняты преимущественно фотосинтетиками и обеспечивают прирост биомассы, нижний образован хемо- и гетеротрофами, минерализующими органику. Эти сложные колонии способны существовать в экстремальных условиях, но не выдерживают конкуренции с более развитыми формами жизни. Проявлением их жизнедеятельности являются карбонатные строматолиты – первая буква палеонтологической летописи.
| Слой | Экогруппа | Автотрофы | Гетеротрофы |
| Верхний. | Фотосинтезирующие аэробы | Цианобактерии: СО2+Н2О+☼→(СН2О)n+О2 | (СН2О)n+О2 → СО2+Н2О+АТФ |
| Промежуточный. | Факультативные аэробы и анаэробы | Пурпурные бактерии: СО2+Н2S+☼→(СН2О)n+S, SO42- | |
| Нижний. | Афотические анаэробные хемотрофы и гетеротрофы | Сульфат-редукторы: SO42-+Н2→ Н2S+Н2О+АТФ. Метаногены: СО2+Н2→СН4+Н2О+АТФ | Бродильщики: (СН2О)n→ СО2+Н2+АТФ |



 Побочным продуктом фотосинтеза был О2. Насыщение атмосферы кислородом происходило также благодаря фотолизу паров воды под воздействием коротковолнового излучения Солнца: Н2О + ☼ ⇒ ОН+ + Н+; 4ОН+ ⇒ О2 + 2Н2О, а также при галогенизации оксидов щелочных и щелочноземельных металлов: 2Na2O + 2С12 ⇒ 4NaCl + О2; 2СаО + 2F2 ⇒ 2CaF2 + О2 и при извержении континентальных вулканов (см.рис.).
Побочным продуктом фотосинтеза был О2. Насыщение атмосферы кислородом происходило также благодаря фотолизу паров воды под воздействием коротковолнового излучения Солнца: Н2О + ☼ ⇒ ОН+ + Н+; 4ОН+ ⇒ О2 + 2Н2О, а также при галогенизации оксидов щелочных и щелочноземельных металлов: 2Na2O + 2С12 ⇒ 4NaCl + О2; 2СаО + 2F2 ⇒ 2CaF2 + О2 и при извержении континентальных вулканов (см.рис.).
Первоначально О2 поглощался при окислении восстановленных минералов (преимущественно металлического железа). Со временем убывание радиогенного тепла и вулканической активности, рост континентов как минеральных ловушек, концентрация химических элементов в пределах биосферы, снижение концентрации СО2 и оксигенизация привели к химическому обеднению гидросферы и атмосферы. Большая часть биофильных тяжелых металлов под действием гравитации переместилась к центру планеты ("железная катастрофа"), часть окислилась, потеряла доступность и подвижность. Приблизительно 600 млн. назад закончилось свободное железо, скорость поступления О2 превысила скорость потребления, и он начал накапливаться в атмосфере.
 Свободный О2, делая одни металлы малодоступными, увеличивал растворимость других, например, благодаря окислению их нерастворимых сульфидов до более растворимых сульфатов. Так, в горячих сероводородных источниках Мо выпадает в осадок в виде сульфида, и живущие там археи-гипертермофилы облигатно зависимы от W. В присутствии О2 Мо становится растворимым, а W недоступным; изменяется и состав микробных сообществ. Во многих филогенетических ветвях Мо заменил W в обменных процессах, и присутствие Ni и W в составе ферментов может рассматриваться в качестве физиологического реликта раннеархейской биосферы (4 млрд. лет назад).
Свободный О2, делая одни металлы малодоступными, увеличивал растворимость других, например, благодаря окислению их нерастворимых сульфидов до более растворимых сульфатов. Так, в горячих сероводородных источниках Мо выпадает в осадок в виде сульфида, и живущие там археи-гипертермофилы облигатно зависимы от W. В присутствии О2 Мо становится растворимым, а W недоступным; изменяется и состав микробных сообществ. Во многих филогенетических ветвях Мо заменил W в обменных процессах, и присутствие Ni и W в составе ферментов может рассматриваться в качестве физиологического реликта раннеархейской биосферы (4 млрд. лет назад).
К концу архея (приблизительно 2 млрд лет назад) концентрация О2 достигла критического значения – 0,2% (основная точка Пастера). На планете разразилась «кислородная катастрофа»[16].
ПРОТЕРОЗОЙ. 2,8 млрд – 600 млн лет назад. Эта эра начинается с кислородной катастрофы, гуронского оледенения и появления эукариот, продолжается формированием и распадом двух суперконтинентов, появлением озонового слоя и многоклеточных водорослей, а заканчивается еще одним оледенением и появлением многоклеточных животных. Активные формы кислорода, образующиеся под воздействием ионизирующего излучения или в процессе жизнедеятельности, являются токсичными – они взаимодействуют с липидами мембран, белками, НК, изменяя их свойства. Поэтому фотосинтетики, вырабатывающие О2, выстроили систему антиоксидантной защиты, которая помогла им пережить кислородную катастрофу, и стали аэробами. Анаэробы нашли другой выход – они стали эукариотами J.
ЭУКАРИОТЫ. С 2,5 до 1 млрд лет до н.э. у прокариот происходит ряд ароморфозов, связанных между собой. На упрощенной схеме они показаны последовательно. Первым переломным моментом в развитии эукариот было формирование цитоскелета – динамичной системы нитчатых белков, в функции которой входит поддержание и адаптация формы клетки ко внешним воздействиям, экзо- и эндоцитоз, обеспечение движения клетки как целого, активный внутриклеточный транспорт и клеточное деление [цит. из Википедии]. Основными компонентами цитоскелета являются 3 системы[17]: 1) кератиновые промежуточные филаменты заякоривают клеточные контакты и поддерживают форму клетки; 2) актин-миозиновая система (микрофиламенты) обеспечивает преимущественно изменение формы мембраны; 3) тубулин-динеиновая система (микротрубочки) обеспечивает внутриклеточную транспортировку молекул и органоидов, слишком крупных, чтобы «плыть по течению» цитоплазмы (в т.ч. и хроматид при делении). Благодаря актин-миозиновой системе становится возможным фагоцитоз – поглощение крупных частиц. Эукариоты стали первыми хищниками, усложнили тем самым трофические цепи, сэкономили энергию на фотосинтезе, заполучили в вечное пользование митохондрии.

| Усложнение синтеза и процессинга белков. Развитие и стабилизация регуляторных (сигнальных) систем. |
| Стабилизация ДНК, появление интронов. Линейные хромосомы. |
| Формирование цитоскелета. |
| Аэробный обмен (дыхание). |
| Совершенствование механизма деления. Митоз и мейоз. |
| Формирование органоидов клетки (в т.ч. ядра), движение и фагоцитоз. |
| Митохондрии, хлоропласты. |
Возможно, основным ароморфозом было совершенствование механизма репарации ДНК, появление гистонов и ядерной оболочки, митоз. Эукариоты научились «выключать» полученные при горизонтальном переносе гены (побочным эффектом стало накопление интронов), бороться с мутациями. Половое размножение (митоз) позволило им избавляться от неудачных мутаций и способствовало комбинативной изменчивости. В целом эволюция размножения идет, как правило, в направлении от бесполых форм к половым, от изогамии к анизогамии, от участия всех клеток в размножении к разделению на соматические и половые, от наружного оплодотворения к внутреннему; к внутриутробному развитию и заботе о потомстве; к уменьшению численности потомства и увеличению его выживаемости; уменьшается роль гаплоидной и увеличивается роль диплоидной фазы. Темп размножения, численность потомства, частота смены поколений наряду с другими факторами определяют скорость приспособления вида к условиям среды. Например, высокие темпы размножения и частая смена поколений позволяют насекомым в короткий срок вырабатывать устойчивость к ядохимикатам. Освоение полового размножения привело к двум важным следствиям. Первым следствием стала самоидентичность, индивидуальность, неповторимость каждой новой комбинации генов, нового организма, и в то же время возможность необратимой потери уникальных аллелей, конечность существования. Живая материя становится дискретной не только в пространстве, но и во времени. Вторым следствием стала репродуктивная изоляция, которая привела к возникновению популяций и видов, к экологической специализац