Трансляция транскрипция
Транскри́пция (от лат. transcriptio — переписывание) — процесс синтеза РНК с использованием ДНК в качестве матрицы, происходящий во всех живых клетках. Другими словами, это перенос генетической информации с ДНК на РНК.
Транскрипция катализируется ферментом ДНК-зависимой РНК-полимеразой. Процесс синтеза РНК протекает в направлении от 5'- к 3'- концу, то есть по матричной цепи ДНК РНК-полимераза движется в направлении 3'->5'[1]
Транскрипция состоит из стадий инициации, элонгации и терминации.
элонгация транскрипции
Момент перехода РНК-полимеразы от инициации транскрипции к элонгации точно не определен. Три основных биохимических события характеризуют этот переход в случае РНК-полимеразыкишечной палочки: отделение сигма-фактора, первая транслокация молекулы фермента вдоль матрицы и сильная стабилизация транскрипционного комплекса, который кроме РНК-полимеразы включает растущую цепь РНК и транскрибируемую ДНК. Эти же явления характерны и для РНК-полимераз эукариот. Переход от инициации к элонгации сопровождается разрывом связей между ферментом, промотором, факторами инициации транскрипции, а в ряде случаев — переходом РНК-полимеразы в состояние компетентности в отношении элонгации (например, фосфорилированиеCTD-домена у РНК-полимеразы II). Фаза элонгации заканчивается после освобождения растущего транскрипта и диссоциации фермента от матрицы (терминация).
На стадии элонгации в ДНК расплетено примерно 18 пар нуклеотидов. Примерно 12 нуклеотидов матричной нити ДНК образует гибридную спираль с растущим концом цепи РНК. По мере движения РНК-полимеразы по матрице впереди нее происходит расплетание, а позади — восстановление двойной спирали ДНК. Одновременно освобождается очередное звено растущей цепи РНК из комплекса с матрицей и РНК-полимеразой. Эти перемещения должны сопровождаться относительным вращением РНК-полимеразы и ДНК. Трудно себе представить, как это может происходить в клетке, особенно при транскрипции хроматина. Поэтому не исключено, что для предотвращения такого вращения двигающуюся по ДНК РНК-полимеразу сопровождаюттопоизомеразы.
: Обратная транскрипция
Некоторые вирусы (такие как ВИЧ, вызывающий СПИД), имеют возможность транскрибировать РНК в ДНК. ВИЧ имеет РНК-геном, который встраивается в ДНК. В результате, ДНК вируса может быть объединено с геномом клетки-хозяина. Главный фермент, ответственный за синтез ДНК из РНК, называется ревертазой. Одной из функций ревертазы является создание комплементарной ДНК (кДНК) из вирусного генома. Ассоциированый фермент рибонуклеаза H расщепляет РНК, а ревертаза синтезирует кДНК из двойной спирали ДНК. кДНК интегрируется в геном клетки-хозяина с помощью интегразы. Результатом является синтез вирусных протеинов клеткой-хозяином, которые образуют новые вирусы. В случае с ВИЧ так же программируется апоптоз (смерть клетки) Т-лимфоцитов.[7] В иных случаях клетка может остаться распростанителем вирусов.
Некоторые клетки эукариотов содержат фермент теломеразу, так же проявляющую активность обратной транскрипции. С её помощью синтезируются повторяющиеся последовательности в ДНК. Теломераза часто активирутся в раковых клетках для бесконечной дупликации генома без потери кодирующей протеины последовательности ДНК.
Трансляция — синтез полипептидной цепи с использованием мРНК в роли матрицы. Как и транскрипция, трансляция — сложный многостадийный процесс, требующий значительных затрат энергии и участия большого числа (до 300) вспомогательных молекул. В трансляции участвуют все три основных типа РНК:
м-, р- и тРНК. мРНК является информационной матрицей; тРНК “подносят” аминокислоты и узнают кодоны мРНК; рРНК вместе с белками образуют рибосомы, которые удерживают мРНК, тРНК и белок и осуществляют синтез полипептидной цепи. Процесс трансляции основывается на том, что каждому триплету мРНК (кодону) соответствует определенная аминокислота. Генетический код расшифровывают (реализуют) тРНК. Напомним, что тРНК имеет структуру, состоящую из четырех петель. К одной из них присоединяется аминокислота (акцепторная петля), в противоположной (антикодоновой) находится триплет нуклеоти-дов, комплементарный кодону мРНК. Этот триплет называют антикодоном. Так, аминокислоте триптофану соответствует кодон УГГ в мРНК, триптофановая тРНК имеет антикодон АЦЦ.
4.хромосомы,их строение,химический состав
Хромосо́мы (др.-греч. χρῶμα — цвет и σῶμα — тело) — нуклеопротеидные структуры в ядре эукариотической клетки, в которых сосредоточена бо́льшая часть наследственной информации и которые предназначены для её хранения, реализации и передачи. Хромосомы чётко различимы в световом микроскопе только в период митотического или мейотического деления клетки. Набор всех хромосом клетки, называемый кариотипом, является видоспецифичнымпризнаком, для которого характерен относительно низкий уровень индивидуальной изменчивости[1].
Исходно термин был предложен для обозначения структур, выявляемых в эукариотических клетках, но в последние десятилетия всё чаще говорят обактериальных или вирусных хромосомах. Поэтому, по мнению Д. Е. Корякова и И. Ф. Жимулёва[2], более широким определением является определение хромосомы как структуры, которая содержит нуклеиновую кислоту и функция которой состоит в хранении, реализации и передаче наследственной информации. Хромосомы эукариот — это ДНК-содержащие структуры в ядре, митохондриях и пластидах. Хромосомы прокариот — это ДНК-содержащие структуры в клетке без ядра. Хромосомы вирусов — это молекула ДНК или РНК в составе капсида.
Морфология метафазных хромосом[править | править исходный текст]
В ходе клеточного цикла облик хромосомы меняется. В интерфазе это очень нежные структуры, занимающие в ядре отдельные хромосомные территории, но не заметные как обособленные образования при визуальном наблюдении. В митозе хромосомы преобразуются в плотно упакованные элементы, способные сопротивляться внешним воздействиям, сохранять свою целостность и форму[5][6]. Именно хромосомы на стадии профазы, метафазы или анафазы митоза доступны для наблюдения с помощью светового микроскопа. Митотические хромосомы можно увидеть у любого организма, клетки которого способны делиться митозом, исключение составляют дрожжи S.cerevisiae, чьи хромосомы слишком малы[7]. Обычно митотические хромосомы имеют размеры в несколько микрон. Например, самая большая хромосома человека хромосома 1 имеет длину около 7 — 8 мкм в метафазе и 10 мкм в профазе митоза[8].
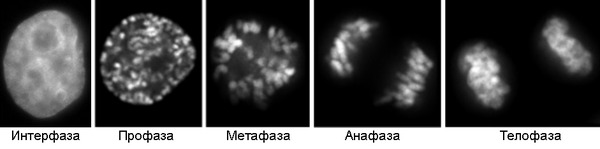

Клетки HeLa в интерфазе и на последовательных стадиях митоза
Метафазные хромосомы состоят из двух продольных копий, которые называются сестринскими хроматидами и которые образуются при репликации. На стадии метафазы сестринские хроматиды соединены в районе первичной перетяжки, называемой центромерой. Центромера отвечает за расхождение сестринских хроматид в дочерние клетки при делении. На центромере происходит сборка кинетохора — сложной белковой структуры, определяющей прикрепление хромосомы к микротрубочкам веретена деления — движителям хромосомы в митозе[9]. Центромера делит хромосомы на две части, называемые плечами. У большинства видов короткое плечо хромосомы обозначается буквой p, длинное плечо — буквой q. Длина хромосомы и положение центромеры являются основными морфологическими признаками метафазных хромосом.
В зависимости от расположения центромеры различают три типа строения хромосом:
· акроцентрические хромосомы, у которых центромера находится практически на конце, и второе плечо настолько мало, что его может быть не видно на цитологических препаратах;
· субметацентрические хромосомы с плечами неравной длины;
· метацентрические хромосомы, у которых центромера расположена посередине или почти посередине[10].
Эту классификацию хромосом на основе соотношения длин плеч предложил в 1912 году российский ботаник и цитолог С. Г. Навашин. Помимо вышеуказанных трёх типов С. Г. Навашин выделял ещё и телоцентрические хромосомы, то есть хромосомы только с одним плечом. Однако по современным представлениям истинно телоцентрических хромосом не бывает. Второе плечо, пусть даже очень короткое и невидимое в обычный микроскоп, всегда присутствует[11].
Дополнительным морфологическим признаком некоторых хромосом является так называемая вторичная перетяжка, которая внешне отличается от первичной отсутствием заметного угла между сегментами хромосомы. Вторичные перетяжки бывают короткими и длинными и локализуются в разных точках по длине хромосомы. Во вторичных перетяжках находятся, как правило, ядрышковые организаторы, содержащие многократные повторы генов, кодирующих рибосомальные РНК. Небольшие хромосомные сегменты, отделяемые от основного тела хромосомы вторичными перетяжками, называются спутниками. Хромосомы, обладающие спутником, принято называть SAT-хромосомами (лат. SAT (Sine Acid Thymonucleinico) — без ДНК).
Необычные типы хромосом
Микрохромосомы
У многих птиц и рептилий хромосомы в кариотипе образуют две чёткие группы: макрохромосомы и микрохромосомы. У некоторых видов микрохромосомы настолько мелкие и их так много, что невозможно отличить одну от другой[18]. Микрохромосомы являются короткими по длине, но обогащёнными генамихромосомами. Например, кариотип курицы содержит 39 хромосом, 6 из которых относятся к макрохромосомам, а 33 — к минихромосомам. Макрохромосомы содержат две трети геномной ДНК, но только 25 % генов, в то время как микрохромосомы содержат оставшуюся треть геномной ДНК и 75 % генов. Таким образом, плотность генов в минихромосомах курицы в шесть раз выше, чем в макрохромосомах[19].
B-хромосомы
B-хромосомы — это добавочные хромосомы, которые имеются в кариотипе только у отдельных особей в популяции. Они часто встречаются у растений, описаны у грибов, насекомых и животных. Некоторые В-хромосомы содержат гены, часто это гены рРНК, однако не ясно, насколько эти гены функциональны. Наличие В-хромосом может влиять на биологические характеристики организмов, особенно у растений, где их наличие ассоциируется с пониженной жизнеспособностью. Предполагается, что В-хромосомы постепенно утрачиваются в соматических клетках в результате нерегулярности их наследования
Голоцентрические хромосомы
Голоцентрические хромосомы не имеют первичной перетяжки, они имеют так называемый диффузный кинетохор, поэтому во время митоза микротрубочки веретена деления прикрепляются по всей длине хромосомы. Во время расхождения хроматид к полюсам деления у голоцентрических хромосом они идут к полюсам параллельно друг другу, в то время как у моноцентрической хромосомы кинетохор опережает остальные части хромосомы, что приводит к характерной V-образной форме расходящихся хроматид на стадии анафазы. При фрагментации хромосом, например, в результате воздействия ионизирующего излучения, фрагменты голоцентрических хромосом расходятся к полюсам упорядоченно, а не содержащие центромеры фрагменты моноцентрических хромосом распределяются между дочерними клетками случайным образом и могут быть утрачены
Голоцентрические хромосомы встречаются у протист, растений и животных. Голоцентрическими хромосомами обладает нематода