Проксимальный извитой каналец (S1). В начальных отделах проксимального извитого канальца (рис. 9-26 А) количество профильтровавшегося HCO3- составляет 90%. Превращение HCO3- в CO2 и ОН- катализирует мембранная карбоангидраза IV IV), расположенная в апикальной мембране. CO2, диффундирует в клетку через люминальную мембрану. В клетке CO2 вступает в реакцию, катализируемую цитоплазматической карбоангидразой II II), ведущую к образованию HCO3-, который частично вступает в реакцию с OH- с образованием CO32- и H2O, таким образом, в клетке содержатся вновь образованные HCO3-, CO32- и H2O. В базолатеральной мембране переносчик Na+-HCO3- (котранспорт) удаляет HCO3- из клетки, но в интерстициальном пространстве HCO3- вступает в реакцию, катализируемую мембранной (CА?) с образованием CO2 и ОН-. В апикальной мембране расположен электронейтральный (обмен 1:1) переносчик NHE3 или, иначе, Na+-H+-переносчик (антипорт), который осуществляет секрецию ионов H+ в просвет канальца в обмен на ионы Na+. Его движущая сила обмена - химический градиент Na+. При его работе ионы H+в результате вторично-активного транспорта секретируются в просвет канальца. Снижение внутриклеточного pH (сдвиг в кислую сторону) активирует данный Na+-H+-переносчик (антипорт), поэтому в случае ацидоза усиленно секретируются ионы H+; и наоборот при алкалозе секреция ионов H+ снижается. Кроме того в апикальной мембране расположена Н+-АТФаза, которая секретирует ионы H+ в просвет канальца. Данный насос позволяет секретировать ионы H+ независимо от реабсорбции Na+. Секреция H+уже в первой трети проксимального канальца снижает рН в просвете канальца с 7,4 (в фильтрате) до 6,5-6,8 (рис. 9-26 А). Эти ионы вступают в реакцию с OH- с образованием H2O, которая уходит в клетку и там формирует ионы H+ и OH-, замыкая, тем самым, цикл.
Проксимальный прямой каналец (S3). Посредством электронейтрального Na+-Н+-переносчика (антипорт), расположенного в апикальной мембране, осуществляется реабсорбция Na+ (рис. 9-26 Б). Кроме того в апикальной мембране расположена Н+-АТФаза, которая секретирует ионы H+ в просвет канальца. Большая часть HCO3- была
реабсорбирована в проксимальном извитом канальце на первой стадии реабсорбции, поэтому H+, выходящий в просвет канальца в результате обмена с Na+ (антипорт), не может быть использован для связывания с HCO3-. Здесь основная роль принадлежит реабсорбции Na+ и ее механизм - обмен Na+/Н+ (NHE3-переносчик). Из клетки в интерстициальное пространство HCO3- попадает двумя путями: в результате обмена С1-/НСO3- (антипорт) и Na+/НСO3- (синпорт).
Дистальный прямой каналец (толстая восходящая часть петли Генле). В этом отделе на апикальной мембране клеток эпителия локализован переносчик, обеспечивающий обмен Na+/H+. Он предотвращает увеличение значения pH канальцевой мочи, которое в верхнем отделе проксимального канальца было снижено до 6,5-6,8, тем более что на этом отрезке нефрона положительный в просвете канальца трансэпителиальный потенциал представляет собой движущую силу для межклеточной реабсорбции H+. Кроме того переносчик, обеспечивающий обмен Na+/H+ (антипорт), расположенный на апикальной мембране, служит для того, чтобы доставлять в просвет канальца ионы H+, возникающие из другого источника: диссоциация реабсорбированного NH4+
на NH3 + H+ (рис. 9-26 В).
Собирательные протоки. В эпителии связующего канальца, коркового и мозгового собирательного протока находятся вставочные клетки α-типа которые имеют в люминальной мембране, как уже упоминалось, Н+/К+-АТФазу и Н+/ АТФазу (рис. 9-26 Г). Вставочным клеткам типа α удается с помощью H+-АТФаз сдвинуть значение рН в собирательном протоке и конечной моче до уровня меньше 5. Образующийся с помощью карбоангидразы HCO3- через базолатеральную мембрану выводится из клетки с помощью переносчика, осуществляющего обмен HCO3- /Cl-, при этом входящий Cl- снова выводится из клетки через Cl--каналы.
Итак, pH в просвете связующего канальца, а также в корковом и мозговом собирательном протоках, может быть значительно сдвинуто в кислую сторону за счет механизма первичного активного транспорта (H+-АТФаза вставочных клеток α-типа), обеспечивающего секрецию ионов H+ в просвет нефрона. При ацидозе оно может достигать значения 4,5. При алкалозе вставочные клетки (вместо ионов H+) выделять бикарбонат в базолатеральной мембране, поэтому значение pH мочи сдвигается до 8,2.
Рис. 9-26. Роль почек в подержании кислотно-щелочного равновесия.
Превращение HCO3- в CO2 (и наоборот) катализируют мембранная (CA IV) и, соответственно, цитоплазматическая (CA II) карбоангидраза клеток канальца

Транспорт NH4+
Аммиак NH3 (соответственно, ионы аммония: NH3 о NH4+) образуется в больших количествах (около 1000 ммоль в день) при расщеплении аминокислот в печени, но он ядовит даже в малых концентрациях, поэтому в печени в среднем около 95% его количества, взаимодействует с эквимолярным количеством HCO3-, и превращается (при потреблении АТФ-) в мочевину, которая выводится с мочой как инертное вещество (2NH4+ + 2HCO3- ↔ NH2 - C(=O) - NH2 + CO2 + 3H2O).
Оставшиеся 5% NH3 ↔ NH4+ (около 50 ммоль в день) попадают в неизменном виде или в виде глутамина в почки, где снова из глутамина образуется NH3 ↔ NH4+, который большей частью выводится из организма.
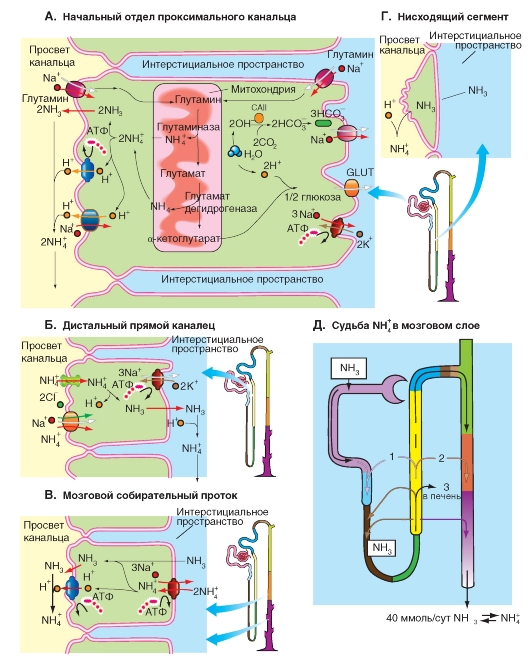
Проксимальный извитой каналец. Глутамин, который переносится с кровью в почки, где он транспортируется в клетки проксимального канальца как через апикальную, так и через базолатеральную мембрану (рис. 9-27 А). В митохондриях этих клеток локализована глутаминаза, которая снова гидролизует глутамин с образованием NH4+ и глутамат-, который посредством глутаматдегидрогеназы расщепляется далее на второй NH4+ и на 2-оксоглутарат2- (а-кетоглутарат2-), при превращении которого в глюкозу используется два иона H+, т.е. образуется два иона HCO3-. Образование этих двух ионов HCO3- остается неучтенным в общем балансе выведения NH4+(↔NH3), поскольку они используются для образования 2-оксоглутарата2-, который в печени необходим для синтеза глутамина (рис. 9-27 А). NH4+ диссоциирует внутриклеточно с образованием NH3 + Н+; оба продукта попадают независимо друг от друга (за счет неионной диффузии и соответственно секреции H+) в просвет канальца, где они вновь образуют NH4+. Последние данные демонстрируют, что по крайней мере такое же количество аммиака выделяется в ионизированной NH4+-форме. В этом случае секреция в просвете канальца осуществляется с помощью Na+-Н+-обменника, который, по всей видимости, вместо ионов H+ может принимать также NH4+.
Небольшие количества NH4+ могут отщепляться от глутамина и в просвете проксимального канальца. Там в качестве «глутаминазы» действует γ-глутамилтрансфераза (γ-GT).
Дистальный прямой каналец (толстая восходящая часть петли Генле).
В результате секреции NH3 и соответственно NH4+ в конце проксимального канальца обнаруживается в 9 раз больше NH4+ (↔ NH3), чем в фильтрате. Однако лишь одна треть этого количества достигает дистального извитого канальца, оставшиеся две трети реабсорбируются (с помощью вторичного активного транспорта) в виде NH4+-ионов в дистальном прямом канальце (рис. 9-27 Б) посредством расположенного на люминальной мембране переносчика BSC1 (котранспорт), который при этом связывает NH4+-ионы вместо K+. После внутриклеточной диссоциации NH4+ (на NH3 и H+) H+ возвращается обратно в просвет канальца (Na+-H+-обменник), а NH3диффундирует из петли Генле в интерстициум мозгового вещества почек, поэтому там устанавливается высокая (возрастающая по направлению к сосочкам до 10 ммоль) концентрация NH4+ о NH3. NH3 попадает оттуда за счет неионной диффузии в просвет собирательного протока.
Кроме того в дистальном прямом канальце на апикальной мембране клеток эпителия локализован переносчик, обеспечивающий обмен Na+/H+. Он предотвращает увеличение значения рН канальцевой мочи, которое в верхнем отделе проксимального канальца было снижено до 6,5-6,8, тем более что на этом отрезке нефрона положительный в просвете канальца трансэпителиальный потенциал представляет собой движущую силу для межклеточной реабсорбции H+.
Просвет собирательного протока
NH3 в просвете собирательного протока (рис. 9-27 В) вследствие обычно очень низкого значения рН тотчас же превращается в NH4+. Почти 80% секретируемого в проксимальном канальце количества попадает в конечную мочу.
На рис. 9-27 Д суммированы пути почечной секреции и выведения аммиака/аммония.
Рис. 9-27. Транспорт аммония (NH4+) - почечная секреция и выведение аммиака/ аммония.
А - на люминальной и базолатеральной мембране клеток проксимального канальца существуют механизмы транспорта (котранспорт с Na+) внутрь клетки глутамина, который образуется в печени и превращается в почках в 2 NH4+ + 2-оксоглутарат2-. NH4+ диссоциирует в клетках канальца на NH3и H+, которые с одной стороны независимо друг от друга попадают в просвет канальца, и с другой стороны в недиссоциированном виде покидают клетку в направлении просвета канальца в качестве заряженных ионов NH4+ с помощью Na+-Н+-переносчика (вместо H+). Б - большая часть образованного в проксимальном канальце NH3 ↔ NH4+реабсорбируется в дистальном прямом канальце (толстой восходящей части петли Генле) в ионизированном виде и попадает в интерстициум мозгового вещества почки. В - из интерстициума мозгового вещества почки соединение в результате неионной диффузии переходит в собирательный проток. Высокая концентрация NH3 ↔ NH4+ в мозговом веществе почек и низкие значения pH мочи в собирательной трубочке ускоряют эту диффузию