Большая часть эффектов лептина проявляется через его воздействие на мозг, куда он проникает через гематоэнцефалический барьер (ГЭБ). Содержание лептина в цереброспинальной жидкости составляет примерно 2-4% от его концентрации в плазме крови, что позволяет предполагать существование специального механизма переноса гормона из крови в ЦНС, который имеет ограниченные возможности. Прохождение лептина через ГЭБ, по всей вероятности, является активным процессом, в котором происходит специфическое связывание лептина с растворимой формой рецептора, выполняющей роль транспортера гормона, и последующим переносом образовавшегося комплекса в мозг (Панков Ю.А., 1998). После такого транспорта через ГЭБ лептин становится доступным для связывания с «длинноцепочечными» рецепторами, наиболее активно экспрессируемыми в гипоталамусе и в меньшей степени – в других структурах мозга.
Клонирование рецептора к лептину и обнаружение его изоформ в мозге сделали «очевидным факт открытия новой физиологической системы регуляции массы тела, в которой лептин играет роль афферентного метаболического сигнала» (Акмаев И.Г., Сергеев В.Г., 2002).
Влияние лептина на пищевое поведение, массу тела и массу жировой ткани опосредуется взаимодействием гормона с нейропептидами и нейромедиаторами, участвующими в регуляции энергетического обмена и продуцируемыми определенными нейронами гипоталамуса, в частности, аркуатного и вентромедиального ядер. Одним из таких нейропептидов является a-меланоцитстимулирующий гормон (a-МСГ), который через меланокортинэргические нейроны ЦНС вызывает снижение аппетита, уменьшение потребления пищи и повышенное использование жиров в энергетическом обмене.
a-МСГ образуется из белкового предшественника – прогормона проопиомеланокортина (ПОМК) в результате его ферментативного расщепления (процессинга). Синтез ПОМК осуществляется в нейронах аркуатного ядра, где установлена экспрессия его гена. В тех же ядрах экспрессируется и ген рецептора лептина. Т.е. эти нейроны несут на своей поверхности рецепторы к лептину, связываясь с которыми, он оказывает на них свое регуляторное влияние. В данном случае это влияние выражается в том, что лептин повышает выраженность экспрессии гена ПОМК, стимулируя тем самым синтез этого прогормона, а через него – и образование a-МСГ (рис. 9.1.4.1).
Еще одним гипоталамическим нейропептидом, с которым взаимодействует лептин, является нейропептид Y. В противоположность a-МСГ он обусловливает повышение аппетита и увеличение потребления пищи. Как следует из рисунка, в отношении данного нейропептида (НPY) лептин проявляет двойной эффект:с одной стороны, тормозит экспрессию гена HPY, снижая тем самым его продукцию, с другой – блокирует физиологические эффекты HPY, предотвращая (или ослабляя) стимулирующее влияние последнего на чувство голода.
Таким образом, лептин реализует свои эффекты на энергетический обмен, проявляющиеся в снижении аппетита, уменьшении потребления пищи, увеличении физической активности и стимуляции сгорания жира в организме. Синергистом лептина в указанных эффектах является инсулин мозговой ткани, который, как и лептин, подавляет экспрессию гена HPY и усиливает секрецию ПОМК. В последнем случае инсулин мозга реализует свое влияние через стимуляцию кортикотропин-рилизинг-гормона (КРГ). Впрочем, такой путь не исключается и для самого лептина, хотя на сегодняшний день он не считается убедительно доказанным.
Лептиновый путь регуляции энергетического гомеостаза не зависит от серотонинового пути, который активируется при приеме таких лекарственных препаратов, как фенфлурамин и дексфенфлурамин (Мantzoros Ch.S., 1999). Таким образом, в организме человека сис –
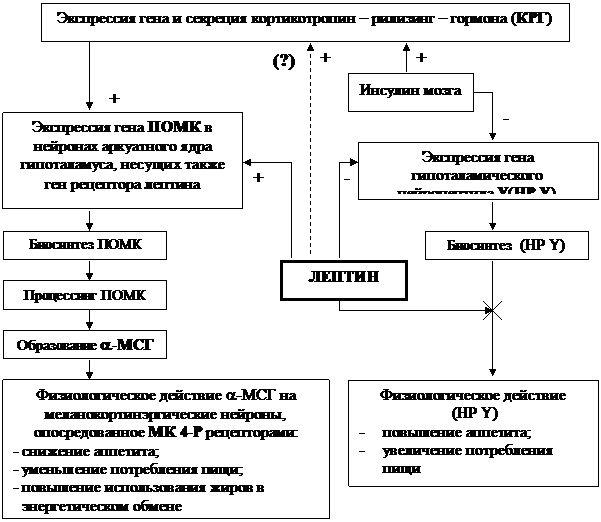 |
Рис. 3 Взаимодействие лептина и нейропептидов гипоталамуса, регулирующих пищевое поведение и энергетический обмен (по Ю.А. Панкову, 1999; 2003).
тема, регулирующая энергетический гомеостаз включает дублирующие друг друга механизмы.
Важно отметить, что в реализации влияния лептина на пищевое поведение существенная роль принадлежит не только (а возможно, не столько) абсолютному содержанию гормона в крови, сколько эффективности его проникновения через ГЭБ. Об этом, в частности, свидетельствуют данные о том, что при анорексии, булимии, неспецифических расстройствах аппетита и депрессии, уровень лептина не отличается от такового у здоровых людей с аналогичной массой тела. Вместе с тем показано, что при нервной анорексии транспорт лептина через ГЭБ находится в обратном соотношении с его концентрацией в крови и наиболее выражен при низком содержании гормонов. Эти данные позволяют объяснить участие лептина в патогенезе нарушений аппетита при нервной анорексии даже в условиях снижения его уровня в циркуляции (Мantzoros Ch.S., 1999).