ОБЩАЯ ФИЗИОЛОГИЯ ЦНС
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ, ПРИНЦИПЫКООРДИНАЦИОННОЙ (ИНТЕГРАТИВНОЙ) ДЕЯТЕЛЬНОСТИ. РЕФЛЕКС КАК ОПЕРАЦИОННЫЙ МЕХАНИЗМ ФУНКЦИОНАЛЬНОЙ СИСТЕМЫ.
Правдивцев В.А. 1, Евсеев А.В. 1, Брук Т.М. 2
1Смоленская государственная медицинская академия
2Смоленская государственная академия физической культуры, спорта и туризма,
Определение понятия «нервный центр»
Нервный центр (НЦ) – объединение нейронов, координирующих и регулирующих всоставефункциональной системы (П. К. Анохин) активность различных физиологических эффекторов, деятельность которых обеспечивает стабильность констант внутренней среды организма (рис. 1). Например, дыхательный центр – объединение нейронов ствола мозга, регулирующих работу дыхательной мускулатуры для поддержания стабильности газовой константы организма; сердечно-сосудистый центр – объединение нейронов продолговатого мозга, регулирующих работу сердца, тонус сосудов для обеспечения стабильности константы оптимального кровяного давления; терморегуляторный центр – нейронный комплекс гипоталамической области, координирующий и регулирующий работу множества эффекторов, активность которых поддерживает постоянство температурной константы организма.
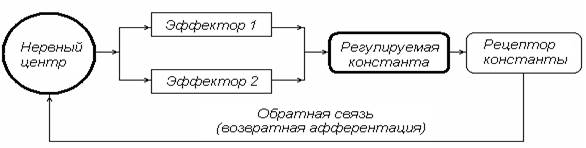
Рис. 1. Принципиальная схема функциональной системы (ФС) организма
Организация нервных центров
Нервные центры вне зависимости от степени сложности организованы практически одинаково. В составе НЦ всегда можно найти входные (афферентные) нейроны, вставочные (промежуточные) нейроны, выходные (эфферентные, моторные) нейроны, аксоны которых связывают НЦ с периферическими эффекторами. Интересно, что в количественном отношении число входных нейронов НЦ обычно превышает число вставочных нейронов, при этом число вставочных нейронов обычно превышает число выходных нейронов. Констатируем, что НЦ организованы по принципу «воронки» (Ч.Шеррингтон). Внутренние связи между нейронами НЦ могут быть самыми разнообразными. В простейшем варианте – это линейные связи.
Элементарной единицей НЦ является нейронныймодуль – совокупность входных, вставочных нейронов, замыкающихся на один выходной (моторный), или конечный нейрон. НЦ обычно состоят из большого числа модулей, функционально связанных друг с другом. На рис. 2 представлена схема простейшего НЦ, в состав которого входят 2 модуля. Каждый отдельно взятый нейронный модуль демонстрирует активность, характерную для всего НЦ.
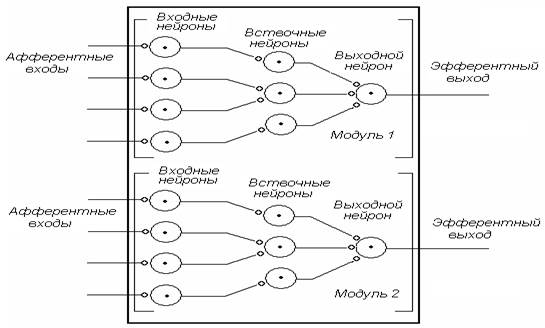
Рис. 2. Принципиальная схема простейшего НЦ, в состав которого входят два модуля (связи между ними – не показаны)
Общие свойства нервных центров
Нервные центры обладают рядом характерных свойств, присущих всем нервным центрам. От чего зависят эти свойства? 1. От свойств одиночных нейронов, входящих в состав НЦ. 2. От свойств химических синапсов, обеспечивающих передачу влияний одних нейронов на другие. 3. От особенностей связей между отдельными нейронами в составе нейронного модуля.
Частные свойства нервных центров
1. НЦ проводит возбуждение только в одну сторону – от входных нейронов к выходным. Данное свойство определяется как односторонняя проводимость. Односторонняя проводимость НЦ объясняется тем, что химические синапсы (рис. 3, синапс 1, синапс 2) проводят возбуждение только в одну сторону – от пресинаптической мембраны к постсинаптической.
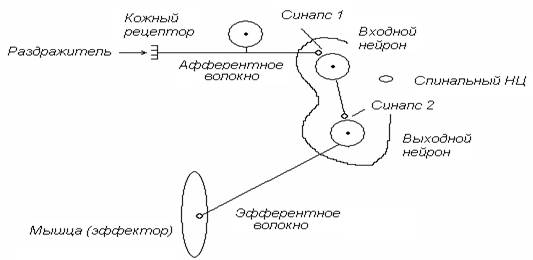
Рис. 3. Схема проведения афферентного возбуждения через спинальный НЦ к скелетной мышце – от входного нейрона к выходному
Одностороннюю проводимость НЦ можно доказать в простом эксперименте с раздражением задних и передних корешков спинного мозга. Если раздражать электрическим током задние корешки, от передних корешков всегда можно зарегистрировать потенциалы действия. Это свидетельствует о проведении возбуждения через НЦ от входных нейронов к выходным. Если раздражать передние корешки, от задних потенциалы действия никогда не регистрируются. Это свидетельствует о том, что от выходных нейронов к входным возбуждение в НЦ не проводится.
2. Возбуждение в нервных центрах распространяется с небольшой скоростью. Данное свойство определяется как замедленное распространение возбуждения.
По периферическим нервным волокнам возбуждение распространяется быстро. Так, по аксонам чувствительных нейронов возбуждение в среднем распространяется со скоростью 50-70 м/с. Однако как только возбуждение поступает в НЦ, скорость его распространения существенно уменьшается, что связано с феноменом синаптической задержки. Время проведения возбуждения через один химический синапс составляет примерно 2 мс (суммарная синаптическая задержка). Следовательно, если возбуждение в НЦ проходит несколько синапсов, средняя скорость проведения возбуждения в НЦ оказывается сниженной по сравнению со скоростью проведения возбуждения в нервных волокнах.
Хорошо известно, что рефлекторные реакции, т. е. изменения активности периферических эффекторов в результате раздражения сенсорных рецепторов и проведения афферентного возбуждения через НЦ, возникают с некоторым скрытым (латентным) периодом, под которым понимаютвремя от момента нанесения раздражения до появления ответной реакции. Значительную часть латентного периода составляет время проведения возбуждения через НЦ. Последнее определяется как центральное время рефлекторной реакции, или рефлекса.
3. Возбуждение в ЦНС может широко иррадиировать от одного НЦ к другим, часто расположенных всамых отдаленных зонах ЦНС.
Иррадиация возбуждения в ЦНС объясняется ветвлением аксонов выходных нейронов нервных центров, связывающих соседние НЦ. При этом создаются возможности для иррадиации возбуждения по ЦНС (рис. 4). Беспрепятственную иррадиацию возбуждения в ЦНС ограничивают тормозные нейроны.
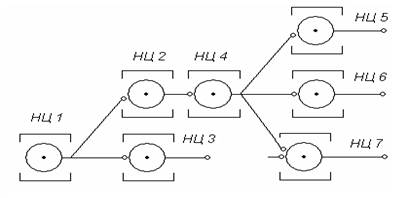
Рис. 4. Иррадиация возбуждения от НЦ1 к другим НЦ (2, 3, 4 и т.д.) в составе ЦНС
Примеры макроорганизации иррадиации периферического возбуждения по коре головного мозга.
Горизонтальная иррадиация. Возбуждение от кожных болевых рецепторов поступает по спиноталамическому тракту, формируя первичный очаг в коре головного мозга. От места возникновения первичного очага возбуждение по горизонтальным ассоциативным связям может широко иррадиировать по коре головного мозга (рис. 5), вовлекая в процесс возбуждения многочисленные корковые нервные центры.
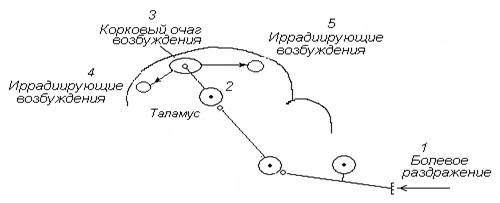
Рис. 5. Механизм горизонтальной иррадиации периферического возбуждения по коре головного мозга
Вертикальная иррадиация. Возбуждение от кожных болевыхрецепторов по спиноталамическому тракту поступает в кору головного мозга, а также по многочисленным коллатералям – в стволовую ретикулярную формацию. По вертикальным ретикуло-кортикальным трактам возбуждение из ретикулярной формации вторично иррадиирует по коре головного мозга, вовлекая в процесс возбуждения многочисленные корковые нервные центры. Данный механизм иррадиации периферического возбуждения по коре головного мозга является основным (рис. 6).
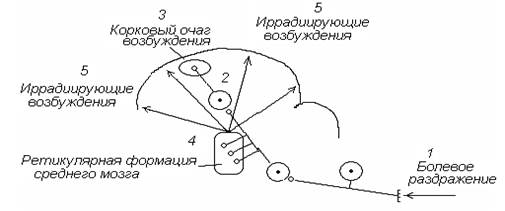
Рис. 6. Механизм вертикальной иррадиации периферического возбуждения по коре головного мозга с участием ретикулярной формации ствола мозга
Возвратная иррадиация. Очаг первичного возбуждения в коре головного мозга по корково-ретикулярным связям может активировать нервные центры восходящей ретикулярной формации ствола мозга, при этом возбуждение из ретикулярной формации может вторично переместиться в кору по вертикальным ретикуло-кортикальным трактам, иррадиируя к различным корковым нервным центрам (рис. 7).
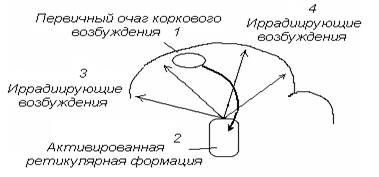
Рис. 7. Механизм возвратной иррадиации возбуждения по коре головного мозга.
4. Нервные центры обладаютсвойством трансформировать, т. е. преобразовывать входной сигнал. Можно указать на 2 типа трансформации. Первый тип – усиление входного сигнала. В этом случае нейронная сеть НЦ работает в режиме «усилителя». Второй тип – ослабление входного сигнала. В этом случае нейронная сеть НЦ работает как «редуктор».
На рис. 8 представлена нейронная сеть НЦ, обеспечивающая режим усиления, или мультипликации (умножения) входного сигнала.
На вход НЦ поступает 2 нервных импульса, на выходе регистрируется 6. Феномен усиления входной импульсации осуществляется в результате ее мультипликации на нейронах 1, 2, 3 НЦ, связанных в конечном итоге с нейроном выходным.
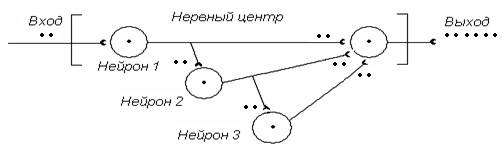
Рис. 8. Схема нейронной сети НЦ, работающей в режиме усилителя входного сигнала в результате его мультипликации на нейронах 1, 2, 3
На рис. 9 представлена нейронная сеть НЦ, обеспечивающая ослабление (редукцию) входного сигнала. Рассмотрим простейшую ситуацию. Линейная цепь нейронов 1-2 не выполняет преобразовательную работу. Введем в нее дополнительный элемент – тормозной нейрон (3), возбуждаемый выходными сигналами клетки 2. Аксон тормозного нейрона соединен с выходным нейроном 2 по схеме возвратного торможения. Что наблюдается на выходе НЦ, если на входной нейрон1 непрерывно будет поступать нервная импульсация?
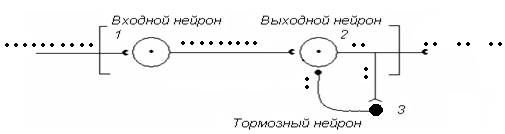
Рис. 9. Нейронная сеть нервного центра с тормозным нейроном, включенным по схеме возвратного торможения
Нейрон 1 принимает поток входных импульсов, передавая их на нейрон 2, который работает в паре с тормозным нейроном 3. От нее возбуждение поступает к нейрону 2, который при этом переходит в состояние торможения. Нейрон 2 перестает реагировать на импульсацию от нейрона 1, следовательно, в структуре выходного потока клетки 2 появляется пауза. Однако, как только клетка 2 перестает работать под влиянием нейрона 3, последний сам перестает работать, т. к. к нему не поступает возбуждающая импульсация. Это немедленно восстанавливает возбудимость нейрона 2, и он вновь начинает реагировать на импульсацию от нейрона 1. Далее циклы повторяются, при этом выходной поток НЦ носит форму, в которой вспышки активности чередуются с тормозными паузами. В целом выходная импульсация оказывается редуцированной в сравнении с исходной.
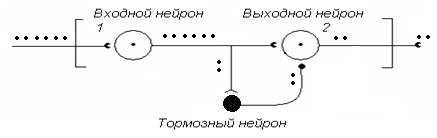
Рис. 10. Нейронная сеть НЦ с тормозным нейроном, включенным по схеме параллельного торможения
Еще один пример с участием тормозного нейрона (рис. 10). Что будет на выходе НЦ, если тормозная клетка включается в нейронную сеть по схеме параллельного торможения?
Под влиянием начальной порции возбуждения входной нейрон 1 генерирует несколько импульсов. Через коллатераль его аксона параллельно активируется тормозной нейрон, который оказывает тормозное влияние на нейрон 2. Его активность блокируется. В итоге – на выходе наблюдаем всего два ПД в сравнении с шестью на входе.
5. Нервным центрам присуще явление последействия, под которым понимают повышенную активность нейронов НЦ на протяжении некоторого времени после прекращения поступления нервных импульсов на вход НЦ. Как этот феномен можно объяснить с позиций нейрофизиологии? Линейная цепь нейронов А-В (рис. 11) последействие не обеспечивает. Однако ситуация изменяется, когда между нейронами А и В встраивается замкнутая сеть вставочных нейронов 1, 2, 3.
Что привносят вставочные нейроны. Итак, от аксона нейрона А коллатераль поступает к нейрону 1, от нейрона 1 аксон поступает к нейрону 2, от нейрона 2 – к нейрону 3, а от нейрона 3 – к входному нейрону А. Формируется «ловушка возбуждения» – циклически замкнутый нейронный комплекс. Возбуждение в таком комплексе может длительно циркулировать, непрерывно поддерживая высокий уровень активности выходного нейрона В на протяжении некоторого времени после прекращения поступления входной импульсации на нейрон А.
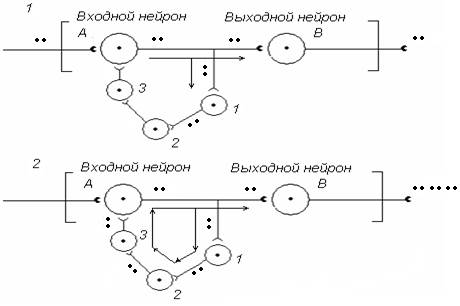
Рис. 11. Схема нейронной сети, обеспечивающей феномен последействия в НЦ за счет циркуляции (реверберации) нервных импульсов в цепи вставочных нейронов 1, 2, 3
Учитывая, что «ловушки возбуждения» в нервных центрах сохраняют рисунок входной импульсации (фактически запоминают параметры входной информации) в настоящее время такого рода объединения нейронов в ЦНС рассматривают в качестве конкретного механизма кратковременной памяти.
6. Нервные центры обладают способностью суммировать возбуждения, поступающие к ним по афферентным каналам. Суммация может быть двух видов: временная и пространственная.
Временная, или частотная суммация (рис. 12). К нейрону НЦ (А) в результате воздействия на входной проводник подходят друг за другом два нервных импульса. Каждый из импульсов вызывает генерацию ВПСП, недостаточную для выведения мембранного потенциала нейрона до критического уровня деполяризации (КУД) – В. Однако если второй импульс приходит к нейрону на фоне развития ВПСП от первого, ВПСП от второго импульса, суммируясь с ВПСП от первого, смещает мембранный потенциал до КУД, нейрон НЦ при этом возбуждается (С). В рассмотренном примере суммация двух входных подпороговых воздействий к НЦ является производным фактора времени между входными воздействиями. При некотором оптимальном временном интервале два подпороговых воздействия могут суммироватьсяна нейронах НЦ, вызывая значимые изменения активности НЦ.
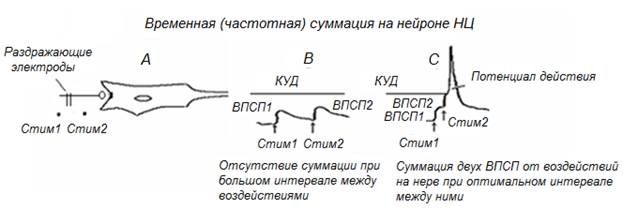
Рис. 12. Временная (частотная) суммация входных воздействий на нейроне НЦ.
Пространственная суммация (рис. 13). К нейрону по различным входным проводникам поступают два нервных импульса (А). Каждый вызывает подпороговый ВПСП (В). Однако если нервные импульсы поступают внейрону НЦ одновременно – подпороговые ВПСП суммируются (С), при этом мембранный потенциал нейрона смещается до КУД, что приводит к возбуждению нейрона НЦ.
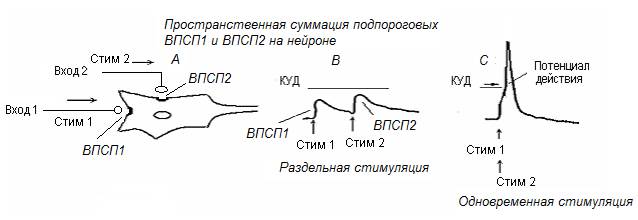
Рис. 13. Пространственная суммация входных воздействий на нейроне НЦ, когда два импульса подходят к нейрону одновременно по различных афферентным проводникам (феномен конвергенции).
7. Нервные центры обладают способностью перестраивать свою работу в зависимости от изменения активности сопряженных нервных центров. Данное свойство определяется как пластичность.
Как доказать феномен пластичности НЦ? На рис. 14-1 представлена упрощенная схема моторных центров спинного мозга А и В, осуществляющих контроль за активностью мышц А и В. Перережем эфферентные (моторные) нервы и соединим их по схеме: нерв А – с мышцей В, нерв В с – мышцей А (рис. 14-2). Спустя некоторое время функциональная целостность нервов восстанавливается, однако моторная активность подопытного животного после вмешательства будет грубо нарушена. Животное теряет способность нормально передвигаться. Тем не менее после многочисленных попыток перемещения в среде обитания ситуация постепенно начинает улучшаться вплоть до приемлемого восстановления моторной активности в результате перестройки нейронной активности спинальных двигательных центров.
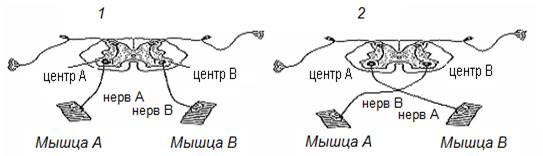
Рис. 14. Упрощенная схема моторных центров спинного мозга интактного животного (1) и животного после перекрестного сшивания эфферентных нервов (2).
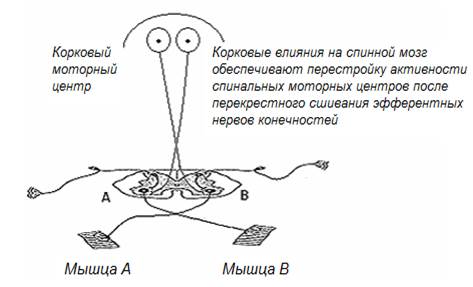
Рис. 15. Упрощенная схема связей коры головного мозга со спинальными центрами
В рассмотренном примере перестройка в работе спинальных нервных центров происходят преимущественно за счет пластических перестроек нейронной активности на уровне коры головного мозга, при этом нисходящие корковые возбуждения нового качества вносят решающий вклад в изменение работы спинальных нервных центров (рис.15). Ведущую роль коры головного мозга в обеспечении пластических перестроек спинальных нервных центров можно доказать в эксперименте с перерезкой нисходящих в спинной мозг корковых двигательных трактов у животного после восстановления двигательных функций. В этой ситуации все немедленно возвращается к исходному состоянию, при котором животное вновь оказывается не способным к перемещению в пространстве.
8.Нервные центры обладают повышенной утомляемостью. Известно, что нервное волокно практически не утомляется (Н. Е. Введенский), напротив, нервные центры обладают повышенной утомляемостью. Неслучайно в целом организме утомление раньше всего развивается в ЦНС (И. М. Сеченов). Причина повышенной утомляемости нервных центров заключается в повышенной утомляемости многочисленных межнейрональных синапсов.
9. Нервные центры обладают повышенной чувствительностью к ядам, биологически активным веществам, лекарственным веществам, что связано с обилием синаптических контактов в нейронных сетях, реализуюших передачу возбуждения от одного нейрона к другому с участием множественных гетерогенных химических механизмов. По этой же причине нервные центры обладают повышенной чувствительностью к недостатку кислорода.
10. Нервные центры могут пребывать в трех функциональных состояниях, отражающих совокупный уровень активности всех нейронов: а) исходное состояние – нормальная (штатная) активность нейронов НЦ; б) сниженная активность нейронов; в) повышенная активность нейронов. В последнем случае НЦ в составе ЦНС становится доминантным, или господствующим (А. А. Ухтомский).
Причины перехода НЦ из исходного состояния в доминантное состояние.
1. Гормональные причины, обычно связанные с избытком или недостатком в циркулирующей крови тех или иных гормонов. 2. Патологические причины. Например, хроническая болевая импульсация, формирующая доминантное состояние центров, ответственных за переработку болевой импульсации (ядра ретикулярной формации, гипоталамуса, таламуса, лимбической системы головного мозга). 3. Метаболические причины. Например, недостаток воды в организме формирует доминантное состояние гипоталамического центра жажды. 4. Информационные причины. Применительно к человеку речь идет о восприятии качества словесных сигналов, письменных и других сообщений, могущих сформировать при определенных ситуациях доминантное состояние корковых центров.
Доминантный НЦ обладает рядом специфических свойств. Среди них – стойкость, повышенная возбудимость нейронов. Повышенная возбудимость доминантного НЦ предопределяет способность нейронов доминантного НЦ реагировать на возбуждения, на которые ранее они не реагировали, иначедоминантный НЦ как бы «притягивает к себе» иррадиирующие по ЦНС возбуждения. На центры-конкуренты доминантный НЦ оказывает угнетающее влияние. Например, человек в состоянии сильной жажды обычно отказывается от еды – доминантный центр жажды угнетает пищевой центр. Доминантный НЦ всегда активирует аппарат биологически отрицательных эмоций, поэтому доминантное состояние того или иного НЦ у человека всегда протекает на фоне неприятных, тягостных эмоциональных ощущений, переживаний. Последнее позволяет сделать вывод о том, что состояниедоминанты в ЦНС формируется как реакция на возникновение жизненно важных проблем, конкретных трудностей и сложностей. Приспособительное значение доминантного состояния НЦ заключается в том, что на основе доминантного НЦ формируется целенаправленный поведенческий акт, ориентированный на достижение таких результатов, которые обеспечивают устранение причин, вызвавших переход соответствующего НЦ из обычного (штатного) состояния в состояние доминантное.
Функцию нервных центров в целом можно определить как координационную, или интегративную. Координационная (интегративная) деятельность нервных центров – деятельность, направленная на обеспечение упорядоченности, согласованности в работе периферических эффекторов, активность которых поддерживает стабильность жизненно важных параметров внутренней среды организма в условиях постоянно изменяющейся среды обитания. Координационная (интегративная) деятельность нервных центров реализуется в составе функциональной системы,под которой понимаютцентрально-периферическую организацию, обеспечивающую достижение полезных для организма конечных результатов. Таким образом, субстратом координационной (интегративной) деятельности НЦ является функциональная система (П. К. Анохин)
В состав ФС (рис. 16) входят: нервный центр; проводники, связывающие НЦ с периферическими эффекторами (канал прямой связи); совокупность периферических эффекторов, определяющих статус регулируемой константы; рецептор регулируемой константы, активность которого отражает ее возможные отклонения от оптимального уровня; канал обратной связи, по которому информация о параметрах регулируемой константы поступает в НЦ, координирующий работу эффекторов, обеспечивая оптимальный для жизнедеятельности организма статус регулируемой константы.
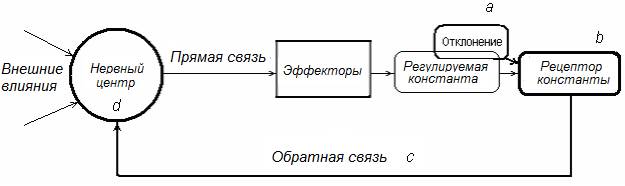
Рис. 16. Схема функциональной системы как субстрата координационной (интегративной) деятельности НЦ – a, b, c, d – последовательность событий в составе функциональной системы, направленных на предотвращение отклонения регулируемой константы от оптимального уровня (описание в тексте).
Возникает вопрос: какой операционный механизм функциональной системы обеспечивает процесс вовлечения НЦ в натуральную координационную (интегративную) деятельность? Ответ: механизм рефлекса, под которым понимают реакцию НЦ, проявляющуюся в изменении физиологической активности эффекторов функциональной системы, под влиянием афферентной импульсации (рис.17) Путь, который проходит нервный импульс в процессе реализации рефлекса, получил название рефлекторной дуги. В состав рефлекторной дуги входят: сенсорный рецептор, афферентные нервы, НЦ (нейроны НЦ), эфферентный нервный проводник, периферический эффектор (эффекторный орган) или периферические эффекторы.
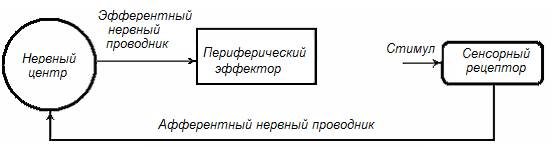
Рис. 17. Принципиальная схема рефлекса как операционного механизма функциональной системы
Примеры простейших рефлекторных актов.
Миотатический рефлекс (коленный рефлекс) – рефлекс в ответ на растяжение мышцы (мышечных веретен), проявляющийся в том, что нейроны спинального НЦ вызывают сокращение этой же мышцы. Схема рефлекторной дуги, включающая всего два нейрона (моносинаптический рефлекс), представлена на рис. 18.
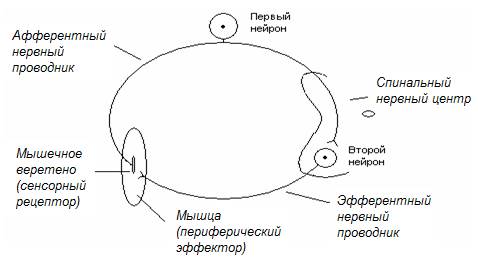
Рис. 18. Схема двухнейронной рефлекторной дуги миотатического рефлекса
Сгибательный рефлекс – рефлекс в ответ на раздражение кожных рецепторов, проявляющийся в том, что нейроны спинального нервного центра вызывают сокращение сгибательных мышц конечности. Схема рефлекторной дуги, включающая три нейрона (полисинаптический рефлекс), представлена на рис. 19.
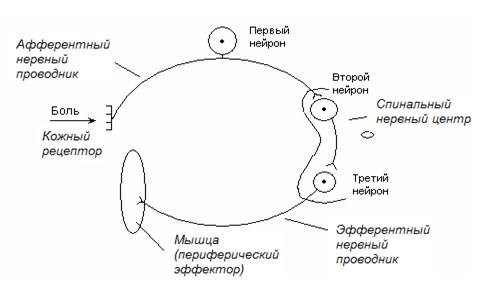
Рис. 19. Схема трехнейронной рефлекторной дуги сгибательного рефлекса