В дополнение к световом и электронно-микроскопических методов, используемых для визуализации acidocalcisomes и описанных выше, спектрофотометрическим методом с использованием Fura-2-паразитов загружены были использованы для обнаружения изменений в acidocalcisome Ca2+(Морено и Zhong, 1996;. Родригес и др, 2002). Acidocalcisomal Ca2+ высвобождается, когда клетки представляются защелачивающие агентов (NH4Cl), ингибиторы VH+-АТФазы (bafilomycin1) И Na+/ЧАС+ (Монензин), и К+/ЧАС+теплообменники, или при инкубации с ионофорами (иономицин) после их подщелачивания (Moreno и Zhong, 1996). Фосфорные соединения были исследованы31Р-ЯМР-спектроскопии (Родригиш и др., 2000;. Moreno и др, 2001), или с помощью биохимических методов (Родригиш и др., 2002). Выделение acidocalci-Somes было сделано с помощью йодиксанол центрифугирования в градиенте с использованием ферментных маркеров (Родригиш и др., 2002). Эта процедура показана на рисунке 10.7. В отличии от процедур, используемых перед тем, чтобы изолировать acidocalcisomes от других клеток, этот метод использовал более низкие концентрации иодиксанола на стадиях градиента, но с образцом добавленного в середине градиента в Йодиксаноле слое 20 процентов, а не прикладывается к вершине градиент без добавления иодиксанола. Этот страт-EGY позволило лучшее разделение acidocalcisomes

258 КАЛЬЦИЯ ХРАНЕНИЕ И гомеостаз токсоплазма
ШЛИФОВАЛЬНЫЕ
ДЛЯ 60S
Тахизоиты лизат переливать


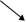
| супернатанте | |||||||||||||||||||||||||
| супернатанте | |||||||||||||||||||||||||
| центрифугирование | |||||||||||||||||||||||||
| PELLET | (36g / 5 мин) | ||||||||||||||||||||||||
| центрифугирование | |||||||||||||||||||||||||
| + | |||||||||||||||||||||||||
| 36g / 5 мин | |||||||||||||||||||||||||
| LYSIS BUFFER | |||||||||||||||||||||||||
| X Ray микроанализа | |||||||||||||||||||||||||
| PELLET | 170 С | ||||||||||||||||||||||||
| супернатанте | |||||||||||||||||||||||||
| Выбрасывания | С | ||||||||||||||||||||||||
| О | |||||||||||||||||||||||||
| PELLET | 10% | U | О | ||||||||||||||||||||||
| центрифугирование | N | ||||||||||||||||||||||||
| п | |||||||||||||||||||||||||
| Выбрасывания | г / 60 мин | T | |||||||||||||||||||||||
| 15% | Калифорния | ||||||||||||||||||||||||
| (144 г / 10 мин) | S | Cu | |||||||||||||||||||||||
| Cu | |||||||||||||||||||||||||
| Mg | |||||||||||||||||||||||||
| PELLET | 20% | 00 | к 100 | ||||||||||||||||||||||
| 25% | |||||||||||||||||||||||||
| супернатанте | TEM | ||||||||||||||||||||||||
| 30% | |||||||||||||||||||||||||
| центрифугирование | супернатанте | ||||||||||||||||||||||||
| 15000 г / 10 мин | Выбрасывания |








Рисунок 10.7 Схематическое представление процедуры фракционирования, используемой для очистки acidocalcisomes Т. гондий тахизоитов. 15 000×г гранулы применяются в среднем слое на 10-30 процентов Йодиксанола ступенчатого градиента. Вставки справа являются Рентгеновским микроанализом изолированной acidocalcisome, показывающего присутствие углерода, кислорода, магния, фосфора и кальция, а также электронно-микроскопического снимок прямой передачи (ПЭЙ) фракции, показывающего электронно-плотных acidocalcisomes. Bar = 1 мкм. Воспроизведено с Permis-Сьон из Родригиш и соавт. (2002), J. Biol. Химреагент 277, 48 650-48 656.
от призраков, присутствующих в промежуточных гидроразрыва-ций и лучшего восстановления фракции осадка у основания градиента (Родригес и др., 2002).
10.5 Са2 + FUNCTION IN
Т. Gondii

Подвижность
Скольжение моторику зависит на актомиозинового двигателя, и имеет важное значение для активного вторжения в клетки-хозяева Т. гондий (Добровольский и Сибли, 1996). В покадровой микроскопии экспериментах паразитов, нагруженных Fluo-4, было обнаружено, что во время паразита
Подвижность цитозольная уровни кальция прошли периодические повышения (как это происходит в мышечных клетках во время схваток), и что внутриклеточные хранит и не внеклеточного кальция были вовлечены в эти изменения (Ловетт и Сибли, 2003).
Коноида экструзия
Коноида экструзии происходит во время вторжения хозяина и выхода из клеток-хозяев, и стимулируется агентами, которые повышают [Са2+]я, Как иономицин, ионофор А23187 кальция и тапсигаргин (Mondragon и Frixione, 1996;. Монтейр и др, 2001). Ионофор-индуцированный коноид экструзия предотвращается путем предварительной инкубации с тахизоитами
| Калифорния2+ Функция в Т. гондий |
внутриклеточный Са2+ хелатор ВАРТА-АМ, но не агентами, которые снижают внеклеточный Ca2+, Как EGTA (Mondragon и Frixione, 1996).