Дарвин и его последователи к основным факторам эволюции относили изменчивость, наследственность и естественный отбор. В настоящее время к ним добавляют множество других дополнительных, неосновных факторов, которые, тем не менее, оказывают влияние на эволюционный процесс, а сами основные факторы понимаются теперь по-новому.
Ведущие факторы эволюции. К ведущим факторам эволюции в настоящее время относят мутационные процессы, популяционные волны численности, изоляцию и естественный отбор.
Поскольку мутации возникают случайно, постольку их результат становится неопределенным, однако случайное изменение становится необходимым, когда оно оказывается полезным для организма, помогает ему выжить в борьбе за существование. Закрепляясь и повторяясь в ряде поколений, случайные изменения вызывают перестройку в структуре живых организмов и их популяций и таким образом приводят к возникновению новых видов. Популяции, насыщенные мутациями, обладают широкими возможностями для совершенствования существующих и выработки новых приспособлений в измененяющихся условиях среды. Однако сам мутационный процесс без участия других факторов эволюции не можетнаправлять изменение природной популяции. Он является лишь поставщиком элементарного эволюционного материала.
Популяционными волнами называют колебания численности особей в популяции. Причины этих колебаний могут быть различными. Например, резкое сокращение численности популяции может произойти вследствие истощения кормовых ресурсов. Среди оставшихся в живых немногочисленных особей могут оказаться редкие генотипы. Если в дальнейшем численность восстановится за счет этих особей, то это приведет к случайному изменению частот генов в генофонде данной популяции. Таким образом, популяционные волны являются поставщиком эволюционного материала.
В качестве третьего основного фактора эволюции СТЭ признает обособленность (изоляцию) группы организмов. На эту особенность указывал еще Дарвин, который считал, что для образования нового вида определенная группа старого вида должна обособиться, но он не мог объяснить необходимость этого требования с точки зрения наследственности. В настоящее время установлено, что обособление и изоляция определенной группы организмов необходимы для того, чтобы она не могла скрещиваться с другими видами и тем самым передавать им и получать от них генетическую информацию. Изоляция разных групп организмов в природе, а также в практике селекционной работы осуществляется разными способами, но цель их одна — исключить обмен генетической информацией с другими видами.
Направляющий фактор СТЭ — естественный отбор. Однако в настоящее время представления о естественном отборе дополнились новыми фактами, значительно расширились и углубились. Естественный отбор следует понимать как избирательное выживание и возможность оставления потомства отдельными особями. Биологическое значение особи, давшей потомство, определяется ее вкладом в генофонд популяции. Отбор действует в популяции, его объектами являются фенотипы отдельных особей. Фенотип организма формируется на основе реализации информации генотипа в определенных условиях среды. Таким образом, отбор из поколения в поколение по фенотипам ведет к отбору генотипов, так как потомкам передаются не признаки, а генные комплексы.
В СТЭ различают три основные формы естественного отбора:
· стабилизирующий,
· движущий
· дизруптивный.
Стабилизирующий отбор способствует сохранению признаков вида в относительно постоянных условиях среды. Он поддерживает средние значения, выбраковывая мутационные отклонения от ранее сформировавшейся нормы. Стабилизирующая форма отбора действует до тех пор, пока сохраняются условия, повлекшие образование того или иного признака или свойства. Примером стабилизирующего отбора является избирательная гибель домовых воробьев при неблагоприятных погодных условиях. У выживших птиц различные признаки оказываются близкими к средним значением. Среди погибших эти признаки сильно варьировались. Примером действия этой формы отбора в популяциях людей служит большая выживаемость детей со средней массой.
Движущий отбор благоприятствует изменению среднего значения признака в измененных условиях среды. Он обусловливает постоянное преобразование приспособлений видов соответственно изменениям условий существования. Особи популяции имеют некоторые отличия по генотипу и фенотипу. При длительном изменении внешней среды, преимущественно в жизнедеятельности и размножении, может появиться часть особей вида с некоторыми отклонениями от средней нормы. Это приведет к изменению генетической структуры, возникновению эволюционно новых приспособлений и перестройке организации вида. Одним из примеров этой формы отбора является потемнение окраски бабочки березовой пяденицы в развитых индустриальных районах Англии. В сельскохозяйственных районах распространены светлоокрашенные формы, а вблизи промышленных центров кора деревьев становится темной из-за исчезновения лишайников, поэтому там преобладает форма темноокрашенных бабочек.
Дизруптивный отбор действует в разнообразных условиях среды, встречающихся на одной территории, и поддерживает несколько фенотипически различных форм за счет особей со средней нормой. Если условия среды настолько изменились, что основная масса вида утрачивает приспособленность, то преимущество приобретают особи с крайними отклонениями от средней нормы. Такие формы быстро размножаются, и на основе одной группы формируется несколько новых. Основной результат этого отбора заключается в наличии нескольких, различающихся по какому-либо признаку групп, как бы разрывающих популяцию. Примером дизруптивного отбора может быть наличие серых и черных волков в Северной Америке, а также существование леопардов обычной и черной окраски (пантер).
Следует отметить, что перечисленные типы отбора очень редко встречаются в чистом виде. Как правило, в живой природе наблюдаются сложные, комплексные типы отбора, и необходимы особые усилия, чтобы выделить из них более простые типы.
Важной составной частью синтетической теории эволюции являются концепции микро- и макроэволюции.
Под микроэволюцией понимают совокупность эволюционных процессов, протекающих в популяциях, приводящих к изменениям генофонда этих популяций и образованию новых видов.Считается, что микроэволюция протекает на основе мутационной изменчивости под контролем естественного отбора. Мутации служат единственным источником появления качественно новых признаков, а естественный отбор — единственным творческим фактором микроэволюции, направляющим элементарные эволюционные изменения по пути формирования адаптации организмов к изменяющимся условиям внешней среды. На характер процессов микроэволюции оказывают влияние колебания численности популяций («волны жизни»), обмен генетической информацией между ними, их изоляция и дрейф генов. Микроэволюция ведет либо к изменению всего генофонда биологического вида как целого, либо к их обособлению от родительского вида в качестве новых форм.
Синтетическая теория эволюции не является застывшей и завершенной концепцией. У нее есть ряд трудностей, на которых основываются недарвиновские концепции эволюции, как уже упоминавшиеся выше, так и недавно возникшие. Так, она допускает возможность изменения геномов организмов в результате мутаций. Но геном любого организма содержит огромное количество нуклеотидов, поэтому мутации не могут повлиять на него так, чтобы получился другой геном. Скорее всего, изменение генома одной клетки или нескольких клеток приведет к рассогласованию в поведении клеток, и популяции клеток не сформируется.
По мнению ряда ученых, приспособленность организмов, естественный отбор и мутации действуют в живой природе, но они не работают в тех масштабах, которые необходимы для образования новых форм.
Так, недавно возникла еще одна концепция недарвиновской эволюции — пунктуализм. Его сторонники считают, что процесс эволюции идет путем редких и быстрых скачков, а в 99% своего времени вид пребывает в стабильном состоянии — стазисе. В предельных случаях скачок к новому виду может совершиться в популяции, состоящей всего из десятка особей, в течение одного или нескольких поколений. Эта гипотеза опирается на широкую генетическую базу, заложенную рядом фундаментальных открытий в молекулярной генетике и биохимии. Пунктуализм отверг генетико-популяционную модель видообразования, идею Дарвина о разновидностях и подвидах как зарождающихся видах и сфокусировал свое внимание на молекулярной генетике особи как носителе всех свойств вида. Ценность этой концепции заключается в идее разобщенности микро- и макроэволюции и независимости управляемых ими факторов.
Возможно, в будущем СТЭ и недарвиновские концепции эволюции, дополняя друг друга, объединятся в новую единую теорию жизни и развития живой природы.
Контрольные вопросы:
1. Что представляет собой вид?
2. Что такое критерии вида? Применения каких критериев достаточно для выделения вида?
3. Какие критерии наиболее объективны для разделения близкородственных видов
4. Что такое ареал? Какие бывают типы ареалов?
5. Что понимают под генетической структурой популяции?
6. Каковы причины непостоянства во времени численности природных популяций?
7. Какие биологические особенности определяют тот или иной тип динамики численности популяции?
8. Какие группы факторов определяют динамику численности популяций?
9. Какие механизмы лежат в основе саморегуляции численности популяции?
10. Раскройте смысл понятий «биологический прогресс» и «биологический регресс»
11. Дайте определение понятиям «ароморфоз», «идиоадаптация» и «общая дегенерация».
12. Приведите примеры ароморфоза
13. Приведите примеры идиоадаптации
14. Приведите примеры общей дегенерации
15. Дайте определения микро- и макроэволюции
Тема 4.4. Макроэволюция
Перечень изучаемых вопросов:
1. Доказательства эволюции.
2. Сохранение биологическогомногообразия как основа устойчивости биосферы и прогрессивного ее развития. Причины вымирания видов.
3. Основные направления эволюционного прогресса. Биологический прогресс и биологический регресс.
Несмотря на невозможность непосредственного исследования макроэволюционных процессов, наука располагает множеством фактов, что эти процессы реальны и существуют в природе.
Палеонтологические данные. Палеонтология как наука оформилась благодаря исследованиям Ж.Кювье (позвоночные), Ж.Б.Ламарка (беспозвоночные) и А. Броньяра (растения). Термин «палеонтология» был предложен французский зоологом Анри Бленвилем (1777—1850) в 1822 г. С помощью палеонтологических методов изучаются ранние этапы развития жизни на Земле, выясняются магистральные пути развития органического мира. Результаты палеонтологических исследований используются для доказательства эволюции животного и растительного мира. Палеонтологические данные и сравнение вымерших форм с современными формами убедительно свидетельствуют о том, что животные и растения постоянно изменялись во времени. Палеонтологам удалось доказать, что одни виды путем длительной эволюции происходят от других исходных форм, и показать это как филогенетические ряды (ряды видов, последовательно сменяющих друг друга в процессе исторического развития).
Филогенетические ряды. По целому ряду групп животных (некоторые копытные, слоны, хищные, моллюски и пр.) палеонтологам удалось воссоздать непрерывные ряды форм (от древнейших до современных), отражающие эволюцию рассматриваемых категорий. Отечественный зоолог В.О.Ковалевский (1842—1883) разработал филогенетический ряд лошадей (рис. 3.13, 3.14): фенакодус (пятипалая конечность) — эогиппус (четырехпалая конечность) — миогиппус (трехпалая конечность) — парагиппус (трехпалая конечность) — плиогиппус (однопалая конечность) — современная лошадь (однопалая конечность). Число пальцев на конечностях уменьшалось в связи с переходом к быстрому и длительному бегу. Одновременно с редукцией числа пальцев увеличивались абсолютные размеры животных (от размера лисицы до современной лошади), осуществился переход от всеядности к исключительной травоядное™, изменилось строение зубной системы и др. На все это ушло до 60 — 70 млн лет. Филогенетические ряды могут рассматриваться как прямые доказательства эволюции.

Переходные формы. Наличие филогенетических рядов не могло служить объяснением для происхождения групп высокого систематического ранга. Противники эволюции жизни на Земле считают возникновение таких групп результатами соответствующих «актов творения». С этой точки зрения особый интерес представляют ископаемые переходные формы, сочетающие в себе одновременно как признаки древних, так и более эволюционно молодых групп, относящихся к таксономическим (систематическим) категориям высокого ранга.
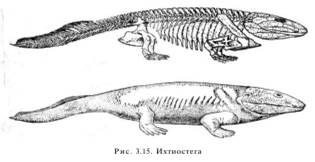

В качестве переходных групп могут быть рассмотрены семенные папоротники (переходная форма между папоротникообразными и голосеменными), ихтиостеги (переходная форма между пресноводными кистеперыми рыбами и земноводными), которых шведский палеонтолог Ярвик образно назвал «четвероногими рыбами» (рис. 3.15).
Данные сравнительной анатомии и внешней морфологии. Не только палеонтологические данные свидетельствуют о наличии переходных форм. В настоящее время также существуют формы, занимающие как бы «промежуточное» положение между категориями высокого систематического ранга. Например, яйцекладущие млекопитающие (утконосы, ехидны) по особенностям своей организации являются промежуточным звеном между рептилиями и млекопитающими.
В строении передних конечностей наземных позвоночных (амфибии, рептилии, птицы и млекопитающие), несмотря на то что все они выполняют самые различные функции, обращает на себя внимание определенное сходство, обусловленное общностью их происхождения. Явление соответствия органов у организмов разных видов, обусловленное их филогенетическим родством, называют гомологией. А такие органы называют гомологичными органами (рис. 3.16). Наличие гомологичных органов у организмов тех или иных групп позволяет проследить степень их родства, определить их филогенетические связи. Но далеко не всякое внешнее сходство в строении органов свидетельствует о родстве организмов. Крыло птицы и крыло бабочки, несмотря на сходство выполняемых функций, а также на некоторое внешнее сходство, по-разному устроены и являются результатом схождения признаков (конвергенции), а не общности происхождения. Такие органы называют аналогичными органами. А явление морфологического сходства органов у организмов различных систематических групп, обусловленное сходством выполняемых ими функций, носит название аналогии. Примеров конвергенции в мире животных много (рис. 3.17—3.19). Колючки некоторых видов растений — аналогичные органы, поскольку их происхождение различно: например колючки боярышника — видоизмененные побеги, колючки кактуса, барбариса — видоизмененные листья.
Доказательствами эволюции являются также рудименты и атавизмы. В организме встречаются недоразвитые органы или их части, не функционирующие у взрослых форм. Это рудименты, или рудиментарные органы (рис. 3.20). По наличию рудиментов можно судить об общности происхождения. Рудименты таза и задних конечностей удавов свидетельствуют о давнем отделении змей от ящериц, а рудименты таза кита — о происхождении от наземных предков, рудименты глаз у слепых пещерных рыб — о том, что эти формы когда-то жили в условиях нормальной освещенности и т.д. Рудименты человека — хвостовые позвонки, ушные мышцы, аппендикс и др. В отличие от рудиментов, которые характерны для всех особей рассматриваемого вида, атавизмы встречаются, как исключение, только у отдельных особей. Атавизмы — это признаки, существовавшие у далеких предков и проявившиеся у отдельных особей вида. К атавистическим структурам следует отнести появление трехпалое™ у современных лошадей, развитие хвоста и волосяного покрова на всем теле у человека и т.д. По атавизмам можно судить, как был устроен тот или иной орган у предковой формы.
Данные эмбриологии. Эмбриология — наука о зародышевом развитии. Исследования, проведенные эмбриологами, показали общность происхождения всех многоклеточных животных, поскольку все они развиваются из одной оплодотворенной яйцеклетки. Все яйцеклетки в своем развитии последовательно проходят определенные стадии (бластулы и гаструлы). Например, все позвоночные животные в процессе эмбрионального (зародышевого) развития проходят стадию закладки жаберных щелей, хотя во взрослом состоянии у наземных позвоночных они отсутствуют. У бескрылых птиц (например, киви) закладываются крылья, у усатых китов на определенных стадиях раннего онтогенеза — зубы, но в дальнейшем они не развиваются.
Самые ранние стадии развития зародышей позвоночных удивительно схожи между собой. В дальнейшем это сходство постепенно утрачивается, все ярче начинают проявляться сначала признаки класса, затем отряда, семейства, рода и, наконец, вида позвоночного животного (рис. 3.21). Таким образом, в процессе индивидуального развития (онтогенеза) каждый вид повторяет свое историческое развитие (филогенез). Выявленная закономерность была сформулирована немецкими биологами Ф. Мюллером (1821 — 1897) и Э.Геккелем (1834—1919) как биогенетический закон, сущность которого заключается в том, что онтогенез есть краткое повторение филогенеза. Естественно, что онтогенез не повторяет абсолютно все этапы филогенеза (некоторые стадии в онтогенезе неотражены). По строению эмбрионов современных форм можно судить о строении эмбрионов (а не взрослых) предков.
Генетические и цитологические данные. Осуществление генетического кодирования, биосинтеза белков и нуклеиновых кислот происходит по единому для всего живого на Земле механизму. Клеточное строение характерно для подавляющего большинства организмов. Принципы деления клеток одинаковы у всех эукариот. Это неоспоримо свидетельствует о едином плане строения и общности происхождения всех организмов. Вероятность того, что принципиально сходная ультраструктура клеток и тончайшие механизмы их функционирования возникли случайно, практически отсутствует и даже теоретически является фантастичной.
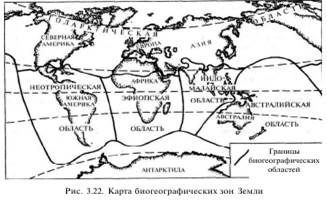
Биогеографические данные. Биогеография изучает закономерности распространения и распределения растений и животных на земном шаре. Своеобразие флоры и фауны Австралии, Новой Зеландии, Южной Америки, Мадагаскара и океанических островов, сходство фауны Северо-Восточной Азии и Северной Америки, Европы и Британских островов, различие фауны Северной и Южной Америки, Африки, Мадагаскара и т.д. — все это свидетельствует о длительной эволюции растительного и животного мира планеты. Причины сходства и различия флоры и фауны различных географических регионов — результат не только исторического развития растительного и животного мира, но и прежде всего тех геологических процессов (дрейф материков, образование островов, появление и исчезновение «сухопутных мостов» между материками, островами и материками и т.д.), которые происходили в это время. А.Уоллес, основываясь на имеющейся в его распоряжении информации о распространении животных, выделил 6 биогеографических областей: Палеарктику, Неоарктику, Эфиопскую, Индомалайскую, Неотропическую, Австралийскую (рис. 3.22).
Сравнение растительного и животного мира различных географических областей позволяет разобраться в их отдаленном прошлом, предоставляет богатый материал, доказывающий эволюцию живых организмов. Сходство фауны и флоры Северной Америки и северо-востока Евразии объясняется наличием в недалеком прошлом узкого перешейка («сухопутного моста») между материками. Такой же мост существовал между Европой и Британскими островами. С другой стороны, длительная (десятки миллионов лет) изоляция Южной Америки (Неотропическая область) от Северной (Неоарктическая область) привела, несмотря на существование в настоящее время Панамского перешейка, к значительным различиям флоры и фауны континентов. Взаимопроникновение видов животных (опоссумы, броненосцы, дикобразы пришли из неотропической области в Северную Америку; олени, медведи, лисы и др. проникли в Неотропики с севера) не изменило характерного (своеобразного) облика фауны Южной и Северной Америки. Еще большим своеобразием отличается животный мир Австралии, которую по праву называют царством сумчатых! Австралия обособилась от Южной Азии более 100 млн лет назад, когда еще не было плацентарных млекопитающих. Разнообразие сумчатых — результат эволюции в условиях длительной географической изоляции.
Особый интерес с точки зрения изучения эволюции представляют острова. Материковые острова (например, Британские острова) имеют фауну и флору, близкую к материковой. Но длительная изоляция острова ведет к ослаблению материковых связей и повышает своеобразие его фауны. Уникальность природы Мадагаскара, ее отличие от природы Африканского континента сложилась в результате длительной географической изоляции острова, отделившегося от материка еще в мезозое. Поэтому на Мадагаскаре нет слонов, жирафов, бегемотов, львов, гиен, леопардов, страусов, зебр, антилоп и прочих представителей фауны Эфиопской области. Высоким процентом эндемичных форм (форм, которые больше нигде не встречаются) характеризуются океанические острова. В целом их видовой состав беден (ущербность фауны и флоры). Например, на таких островах отсутствуют наземные млекопитающие, амфибии, т.е. целые группы видов, не способные преодолевать значительные водные пространства. Для фауны и флоры океанических островов большое значение имеет явление случайного заноса животных (птицы, насекомые, рептилии, последние могут преодолевать водные преграды на стволах деревьев) и семян растений. На островах процесс эволюции в условиях их длительной географической изоляции, в отсутствие генетических связей с материнскими популяциями, формируется эндемичная флора и фауна. Так, на Галапагосских островах эндемизм среди птиц достигает почти 80% (82 вида из 108). А знаменитые галапагосские, или дарвиновы, вьюрки (14 видов) стали первым примером влияния изоляции на видообразование. Ч.Дарвин обратил пристальное внимание на строение клювов у различных видов вьюрков и, проведя исследования, пришел к заключению, что формы клювов сформировались в зависимости от состава поедаемых кормов и способов их добывания. Исследования ученых последующих поколений (Д.Лэк, Э.Майр) показали, что все дарвиновы вьюрки произошли от одного общего материкового предка и случилось все это в результате процесса адаптивной радиации, т.е. в результате расхождения (дивергенции) форм от исходной предковой в ходе приспособительной эволюции. Эффект адаптивной радиации также наглядно демонстрируется на гавайских цветочницах — эндемичной группе птиц, обитающей на Гавайских островах. Известно около 40 видов этих птиц, включая вымершие формы.