РЕГЕНЕРАЦИЯ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫПОЗВОНОЧНЫХ
Восстановление поврежденных аксонов
Шванновские клетки периферической нервной системы обеспечивают окружение, способствующее регенерации аксона. Стимулирующая рост нейронов активность шванновских клеток является следствием секреции многих трофических факторов, экспрессии на поверхности клеток молекул адгезии и интегринов, а также продукции компонентов внеклеточного матрикса, таких как ламинин. Например, эксперименты, в которых повреждался седалищный нерв, показали, что в то время, как периферический участок аксона дегенерирует, уцелевшие шванновские клетки в этой области синтезируют в высоких концентрациях два нейротрофических фактора: BDNF (нейротрофический фактор, полученный из мозга) и NGF. Таким образом, шванновские клетки могут снабжать трофическими факторами BDNF и NGF двигательные, чувствительные и симпатические аксоны, регенерирующие к своим периферическим мишеням. Интересно, что такие «денервированные» шванновские клетки экспрессируют на своей поверхности большое число низкоаффинных рецепторов для NGF и BDNF. Возможно, это поддерживает некоторую тоническую концентрацию этих нейротрофинов, которые служат для выбора правильного направлении для роста регенерирующих аксонов84). После регенерации нервов шванновские клетки прекращают продукцию NGF и BDNF и вновь обеспечивают изоляцию аксонов.
Аполипопротеин E (ApoE), синтезируемый шванновскими клетками и макрофагами, также накапливается в дистальном участке поврежденных периферических нервов и связывается с базальной мембраной шванновских клеток (см. рис. 2). АроЕ поддерживает жизнеспособность нейронов за счет защиты клеток от окислительного повреждения, а также стимуляции роста и адгезии клеток.
 Рис.1. Спраутинг аксонов в ЦНС млекопитающих. (А) Типичная гранулярная клетка зубчатой фасции на ее дендритах во внешнем молекулярном слое (OML) получает множественные сина птические входы от ипсилатеральной энторинальной коры головного мозга и единичные входы от контралатеральной коры и медиальной перегородки. Дендриты внутреннего молекулярного слоя (IML) получают входы от медиальной перегородки и коммиссуральных/ассоциативных волокон. GCL= гранулярный клеточный слой. (В) Удаление ипсилатеральной энторинальной коры приводит к интенсивному росту аксонов нервных клеток энторинальной коры с контралатеральной стороны и замещению ипсилатерального энторинального входа в OML. Наблюдается также спраутинг аксонов, приходящих в DML от медиальной перегородки. Коммиссуральные/ассоциативные волокна в IML расширяют зону иннервации. (С, D) Примеры терминальных разветвлений аксонов от контралатеральной энторинальной коры в OML зубчатой фасции. (С) Норма. (D) Два месяца спустя после повреждения ипсилатеральной энторинальной коры.
Рис.1. Спраутинг аксонов в ЦНС млекопитающих. (А) Типичная гранулярная клетка зубчатой фасции на ее дендритах во внешнем молекулярном слое (OML) получает множественные сина птические входы от ипсилатеральной энторинальной коры головного мозга и единичные входы от контралатеральной коры и медиальной перегородки. Дендриты внутреннего молекулярного слоя (IML) получают входы от медиальной перегородки и коммиссуральных/ассоциативных волокон. GCL= гранулярный клеточный слой. (В) Удаление ипсилатеральной энторинальной коры приводит к интенсивному росту аксонов нервных клеток энторинальной коры с контралатеральной стороны и замещению ипсилатерального энторинального входа в OML. Наблюдается также спраутинг аксонов, приходящих в DML от медиальной перегородки. Коммиссуральные/ассоциативные волокна в IML расширяют зону иннервации. (С, D) Примеры терминальных разветвлений аксонов от контралатеральной энторинальной коры в OML зубчатой фасции. (С) Норма. (D) Два месяца спустя после повреждения ипсилатеральной энторинальной коры.
|
Тем не менее, у нокаутных мышей, лишенных АроЕ, регенерация и ремиелинизация периферических аксонов существенно не меняется. Однако мыши с отсутствием АроЕ имеют сниженное число немиелинизированных аксонов и, соответственно, сниженную чувствительность к повреждающим тепловым стимулам.
При повреждении периферического нерва в зоне повреждения активируются факторы, стимулирующие пролиферацию шванновских клеток. Они включают два цитокина: фактор, ингибирующий лейкемию (ФИЛ), и Reg-2, являющийся потенциальным митогеном шванновских клеток (см. рис. 2). Reg-2 специфически экспрессируется в развивающихся и регенерирующих двигательных и чувствительных нейронах, и эта экспрессия усиливается ФИЛ. Антитела к Reg-2 замедляют регенерацию после передавливания седалищного нерва.
Специфичность реиннервации
Для полного восстановления функции после повреждения нервной системы регенерирующие аксоны должны восстановить связи со своими прежними мишенями. Классические эксперименты Лэнгли, подтвержденные позже исследованиями одиночных клеток, показали, что регенерирующие преганглионарные аксоны нейронов автономной нервной системы реиннервируют соответствующие постганглионарные нейроны. В этом механизме имеет значение, по-видимому, сигнал о позиции нейрона, который оказывает влияние на формирование синапса между нейроном и клеткой-мишенью.
| Рис. 2. Шванновские клетки способствуют регенерации аксонов в периферической нервной системе позвоночных. После аксотомии дистальный участок аксона и миелиновая оболочка дегенерируют и фагоцитируются. Пролиферация шванновских клеток стимулируется двумя цитокинами: фактором, ингибирующим лейкемию (LIF), из макрофагов и Reg-2 из окончаний аксонов. Экспрессия Reg-2 усиливается LIF. Пролиферирующие шваннов ские клетки синтезируют два нейротрофических фактора, BDNF и NGF, которые сохраняются на клеточной поверхности их низкоаффинными рецепторами. Нейротрофины поддерживают процесс регенерации аксонов и направляют их к соответствующим мишеням. Шванновские клетки и макрофаги синтезируют также аполипопротеин Ε (ΑροΕ), который способствует выживанию нейронов и регенерации их аксонов. | 
|
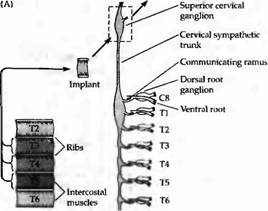
Этот механизм справедлив как для симпатической, так и двигательной системы. Так, если межреберная мышца, извлеченная из передней части грудной клетки, трансплантируется в область шеи, то она преимущественно реиннервируется аксонами шейного симпатического ствола, расположенными в самых передних участках спинного мозга (рис. 3). Наоборот, мышцы, трансплантированные из задних областей тела, имеют тенденцию к реиннервации аксонами более дорзальньгх нейронов. Подобная тенденция установлена и для трансплантации симпатических ганглиев различного исходного расположения. У молодых крыс зависимость от позиции нейрона показана в реиннервации мышц, имеющих множественную иннервацию несколькими сегментами.
Нервно-мышечные синапсы у новорожденных крыс, головастиков и взрослых тритонов после поперечного пересечения двигательного нерва с высокой точностью воссоединяются с исходной синаптической зоной. Одним из механизмов избирательного восстановления синапсов является конкуренция между аксонами. В мышцах саламандры, иннервированных чужим аксоном, чужеродный синапс элиминируется после того, как нормальный нерв восстановил свой синаптический контакт. У взрослых млекопитающих чувствительные, двигательные и постганглионарные аксоны показывают более низкую избирательность по отношению к их исходным мишеням. Чужие нервы при иннервации мышечных волокон могут быть столь же эффективными, как и оригинальные нервные волокна. Более того, чужой нерв может даже вытеснить исходный аксон в интактных мышцах взрослой крысы). Избирательная регенерация у взрослых млекопитающих возможна, если периферический нерв был не перерезан, а передавлен так, чтобы сохранились эндоневрий и базальная мембрана шванновских клеток, окружавших интактные аксоны (см. рис. 4)). При таких условиях регенерирующие аксоны направляются вновь к своим исходным мишеням. Если эндоневрий был разрушен, например при перерезке нерва, регенерация происходит беспорядочно и аксоны часто создают синапсы с неподходящими мишенями.
|
| Рис. 3. Селективная реиннервация мышц, основанная на сегментарном происхождении. (А) Межреберная мышца из грудного сегмента ТЗ, Т4 или Т5 была трансплантирована на шею взрослой крысы и после удаления верхнего шейного ганглия была реиннервирована преганглионарными аксонами шейного симпатического ствола. Сегментарное происхождение входов было определено регистрацией активности мышц при стимуляции отдельных вентральных корешков, которые соединяют преганглиальные аксоны с нервным стволом. (В) Распределение входов к трансплантированным ТЗ, Т4 и Т5 мышцам. Передние мышцы склонны к реиннервации передними преганглиальными аксонами. |