Генетический портрет десяти малых народов Южной Сибири. Сообщение III. Генетические соотношения и положение среди народов Северной Азии по данным об аутосомных ДНК маркерах
Лавряшина М.Б.1, Ульянова М.В.1, Балаганская О.А.2, Балаганский А.Г.3, Кузнецова М.А.3, Балановская Е.В.2
1 ГОУ ВПО Кемеровский Государственный Университет,
650043, г. Кемерово, ул. Красная, д.6, факс 8(3842)583885, lmb2001@mail.ru
2 ГУ Медико-генетический научный центр РАМН, 115478, г. Москва, ул. Москворечье, д.1, факс 007(095)3240702, balanovska@mail.ru
3ГОУ ВПО Читинский Государственный Университет, 672039, г. Чита, ул. Александро-Заводская, 30, bal_ag@mail.ru
АННОТАЦИЯ
По данным о полиморфизме девяти аутосомных ДНК маркеров (SNP: CCR2 Val64Ile; INS/DEL: CCR5del32; Alu: ACE1, TPA25, A25, PV92, ApoA1, B65, F13B) получены оценки генетических расстояний (d) между генофондам десяти малых народов Южной Сибири (суммарная выборка – 1002 человека). Северные алтайцы (N=172) представлены кумандинцами, тубаларами и челканцами; южные алтайцы (N=284) – алтай-кижи и теленгитами; хакасы (N=349) – качинцами, койбалами, кызыльцами и сагайцами; шорцы (N=197) – абаканской и горной территориальными группами. Генетические дистанции (0.0033 ‹ d ‹ 0.0477) отражают общность этногенеза южносибирских народов и их сохраняющуюся генетическую гетерогенность. Наибольшее сходство обнаружено между: алтай-кижи и теленгитами (d =0.003); тубаларами и челканцами (d =0.005); качинцами и койбалами (d =0.008); шорцами абаканскими и горными (d =0.005). Отметим неожиданно большие генетические различия между хакасскими народами. Сагайцы и качинцы сближаются с северными алтайцами, а кызыльцы и койбалы – наиболее монголоидные из хакасских народов – объединяются в один кластер с южными алтайцами, для которых также характерна большая выраженность монголоидных черт. Все это ставит под сомнение правомерность объединения этнических хакасских групп в одно этнотерриториальное объединение при молекулярно-генетических исследованиях.
Аутосомные ДНК маркеры, генетические расстояния, коренные народы Сибири
ВВЕДЕНИЕ
В настоящее время проведен значительный объем исследований, посвященных положению генофондов отдельных народов Сибири и Северной Азии в общей системе евразийского генофонда [5, 7, 15, 19, 22, 23, 25 и многие другие]. Благодаря этим работам на генетической карте Северной Евразии остается все меньше «белых пятен». Однако население контактных зон, к которым относится и Южная Сибирь, является крайне проблематичным объектом для популяционно-генетических исследований. «Саяно-Алтайский этногенетический узел» [1] – термин, достаточно емко характеризующий сложность этнической истории коренного населения Южной Сибири. Разнообразие антропологических типов современных народов Сибири [2, 6, 13, 21] сформировалось в результате разнонаправленных процессов. С одной стороны, это смешение народов с разной расовой и языковой принадлежностью, а с другой – рост подразделенности вследствие географической изоляции, культурных, религиозных традиций и т.д. В итоге возникла антропологически и генетически разнородная общность сибирских тюрок, причем у многих из них до недавнего времени отсутствовали устойчивые самоназвания. Современные этнические классификации коренного населения Сибири в силу объективных причин оказались несовершенны, а этнонимы, которые эти народы используют в качестве самоназваний, зачастую имеют экзогенную природу: были искусственно созданы в связи с научными классификациями народов региона или же возникли под влиянием административной политики [20].
Сложность изучения сибирских этносов требует проведения специальных исследований их генетической структуры и генетического сходства на разных уровнях популяционной иерархии. В настоящей статье акцент сделан не на традиционном анализе сходства генофондов алтайцев, хакасов и шорцев, а на соотношении генофондов малых народов, входящих в их состав. Алтайцы представлены северными алтайцами (кумандинцами, тубаларами и челканцами) и южными алтайцами (алтай-кижи и теленгитами); хакасы – качинцами, койбалами, кызыльцами и сагайцами; шорцы – горной и абаканской территориальными группами.
МАТЕРИАЛЫИ МЕТОДЫ
Материал для исследования был собран в ходе комплексных экспедиций 2003-2009 гг. в Южную Сибирь при сотрудничестве кафедры генетики Кемеровского государственного университета и лаборатории популяционной генетики человека Медико-генетического научного центра РАМН (г. Москва). Осуществлено обследование генофондов коренного населения тринадцати административных районов четырех сибирских регионов: Алтайского края (Красногорский, Солтонский районы); Кемеровской области (Таштагольский район); Республики Алтай (Кош-Агачский, Онгудайский, Турочакский, Улаганский, Чойский районы); Республики Хакасия (Аскизский, Бейский, Орджоникидзевский, Таштыпский, Ширинский районы). Принципы сбора первичных данных, формирования выборок, а также описание методов генотипирования и результатов анализа полиморфизма, уровня гетерозиготности и степени подразделенности анализируемых популяций южносибирских народов приведены в предыдущих сообщениях [9, 10]. Совокупная выборка представителей коренных южносибирских народов составила 1002 неродственных индивидуума.
Для сравнительного описания генофонда алтайцев, хакасов и шорцев были выбраны аутосомные ДНК маркеры, по которым наиболее полно охарактеризованы соседние этносы, имеющие общие этногенетические корни с исследуемыми народами Южной Сибири: SNP CCR2 Val64Ile; INS/DEL CCR5del32; Alu: ACE1, TPA25, A25, PV92, ApoA1, В65, F13B. На основе частот аллелей определены матрицы генетических расстояний (d) [24]. Для визуализации генетических взаимоотношений между группами в программе Statistica методами Уорда и средней связи построены дендрограммы, давшие сходные результаты. Оценка положения генофонда изученных южносибирских этносов в структуре генофондов народов Северной Евразии проведена также методом многомерного шкалирования. Для этого были привлечены литературные данные об аллельных частотах восьми аутосомных ДНК маркеров: CCR2 Val64Ile, CCR5del32, Alu: ACE1, TPA25, A25, PV92, ApoA1, F13B у других евразийских народов (см. базу данных на сайте http//:www.genofond.ru). Для оценки качества результатов шкалирования применяли показатель стресса (SO) Д. Краскела и коэффициент отчуждения – алиенации (КО) Л. Гуттмана. Результаты шкалирования считали приемлемыми при величинах SO и КО, не превышающих 0.10 – 0.15 [8].
РЕЗУЛЬТАТЫИ ОБСУЖДЕНИЕ
Долгая история межэтнических контактов и общие этногенетические корни алтае-саянских народов до сих пор находят отражение в определенной общности их физического облика, языка, культурных традиций и родовой структуры. Генетические расстояния по 9 ДНК маркерам (таблица) между изученными народами Южной Сибири невелики (0.0033 ‹ d ‹ 0.0477) по сравнению со средней дистанцированностью от них татар (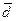 = 0.072), русских ( = 0.061), белорусов ( = 0.074) и украинцев ( = 0.069), но значительно больше, чем, например, различия между народами восточнославянской группы (0.0031< d <0.0034). Это, с одной стороны, отражает единство происхождения южносибирских тюрок и согласуется с данными антропологии, свидетельствующими об относительной расовой однородности основной части коренного населения Хакасии, Шории и Северного Алтая [13]. А с другой стороны говорит о значительной генетической гетерогенности этносов Южной Сибири. Как нами было показано ранее [10], южносибирские народы сохраняют генетическую подразделенность. Для того чтобы выявить степень генетического сходства и различия десяти малых народов Южной Сибири, их анализ проведен в трех аспектах: 1) соотношение генофондов в пределах этнотерриториальных групп (алтайцы, шорцы, хакасы); 2) соотношение генофондов безотносительно к территориальным группам; 3) положение в системе народов Евразии.
= 0.072), русских ( = 0.061), белорусов ( = 0.074) и украинцев ( = 0.069), но значительно больше, чем, например, различия между народами восточнославянской группы (0.0031< d <0.0034). Это, с одной стороны, отражает единство происхождения южносибирских тюрок и согласуется с данными антропологии, свидетельствующими об относительной расовой однородности основной части коренного населения Хакасии, Шории и Северного Алтая [13]. А с другой стороны говорит о значительной генетической гетерогенности этносов Южной Сибири. Как нами было показано ранее [10], южносибирские народы сохраняют генетическую подразделенность. Для того чтобы выявить степень генетического сходства и различия десяти малых народов Южной Сибири, их анализ проведен в трех аспектах: 1) соотношение генофондов в пределах этнотерриториальных групп (алтайцы, шорцы, хакасы); 2) соотношение генофондов безотносительно к территориальным группам; 3) положение в системе народов Евразии.
Таблица
Матрица генетических расстояний (d) между десятью малыми южносибирскими народами (9 локусов)
| Территориальные группы | АЛТАЙЦЫ(N=456) | ХАКАСЫ(N=349) | ШОРЦЫ(N=197) | |||||||||
| Южные (N=284) | Северные (N=172) | |||||||||||
| Этносы | алтай-кижи (N=90) | теленгиты (N=194) | кумандинцы (N=15) | тубалары (N=86) | челканцы (N=71) | качинцы (N=122) | койбалы (N=51) | кызыльцы (N=15) | сагайцы (N=157) | абаканские (N=38) | горные (N=158) | |
| алтай-кижи | .0033 | .0269 | .0198 | .0244 | .0087 | .0071 | .0130 | .0235 | .0065 | .0122 | ||
| теленгиты | .0033 | .0234 | .0188 | .0245 | .0092 | .0034 | .0095 | .0263 | .0125 | .0221 | ||
| кумандинцы | .0268 | .0234 | .0167 | .0250 | .0209 | .0275 | .0140 | .0477 | .0392 | .0439 | ||
| тубалары | .0198 | .0188 | .0167 | .0053 | .0056 | .0146 | .0120 | .0209 | .0191 | .0198 | ||
| челканцы | .0244 | .0245 | .0250 | .0053 | .0054 | .0193 | .0227 | .0176 | .0209 | .0217 | ||
| качинцы | .0087 | .0092 | .0209 | .0056 | .0054 | .0078 | .0121 | .0186 | .0087 | .0112 | ||
| койбалы | .0071 | .0034 | .0275 | .0146 | .0193 | .0078 | .0077 | .0189 | .0091 | .0188 | ||
| кызыльцы | .0130 | .0095 | .0140 | .0120 | .0227 | .0121 | .0077 | .0321 | .0145 | .0234 | ||
| сагайцы | .0235 | .0263 | .0477 | .0209 | .0176 | .0186 | .0189 | .0321 | .0214 | .0197 | ||
| шорцы абаканские | .0065 | .0125 | .0392 | .0191 | .0209 | .0087 | .0091 | .0145 | .0214 | .0045 | ||
| шорцы горные | .0122 | .0221 | .0439 | .0198 | .0217 | .0112 | .0188 | .0234 | .0197 | .0045 | ||
ГЕНЕТИЧЕСКИЕ ВЗАИМООТНОШЕНИЯ МАЛЫХ НАРОДОВ ЮЖНОЙ СИБИРИ В ПРЕДЕЛАХ ЭТНОТЕРРИТОРИАЛЬНЫХ ОБЪЕДИНЕНИЙ
Алтайцы. Построенная на основе матрицы генетических расстояний дендрограмма (рис. 1) выявила два кластера – южноалтайский и североалтайский, что подтверждает дифференциацию коренного населения Алтая на северную и южную подгруппы, ранее показанную совокупностью этноисторических и лингвистических исследований [14], данных антропологии (соматологии, одонтологии и краниологии) [6, 21] и этнической генетики [12].
В пределах каждой из этих общностей наблюдаются минимальные оценки генетических расстояний – внутри южноалтайской (алтай-кижи и теленгиты, d = 0.0033) и североалтайской (тубалары и челканцы d = 0.0053). Кумандинцы, также относящиеся к северным алтайцам, несколько дистанцируются и от тубаларов (d = 0.0167), и от челканцев (d = 0.0250), хотя и входят в единый с ними кластер (см. рис. 1). Полученный результат согласуется с мнением антропологов и этнографов [18] об «особом» положении кумандинцев в составе северных алтайцев. Кроме того, величина показателя средней удаленности от всех южносибирских популяций среди алтайцев так же наиболее высока именно у кумандинцев (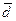 = 0.024). Такая генетическая «удаленность» кумандинцев, подтвержденная и результатом двухмерного шкалирования (рис. 3), может быть следствием их значительной метисированности. Уже в начале ХХ века А.И. Ярхо отмечал сильную смешанность кумандинцев с русскими [21]. Высокая частота браков с русскими свойственна данному этносу и в настоящее время, о чем свидетельствует данные литературы [17], и наши собственные исследования [11]. Метисированность и дисперсность расселения по территории России послужили главной причиной малого объема собранной выборки кумандинцев (15 чел.), в которую включены только те индивиды, у которых все предки на протяжении трех поколений являлись кумандинцами. Тем не менее, необходимо отметить, что хорошая сопоставимость наших данных о генофонде кумандинцев с данными антропологии и этнографии свидетельствует о том, что исследование этого алтайского народа проведено хоть и на малой, но хорошо собранной выборке.
= 0.024). Такая генетическая «удаленность» кумандинцев, подтвержденная и результатом двухмерного шкалирования (рис. 3), может быть следствием их значительной метисированности. Уже в начале ХХ века А.И. Ярхо отмечал сильную смешанность кумандинцев с русскими [21]. Высокая частота браков с русскими свойственна данному этносу и в настоящее время, о чем свидетельствует данные литературы [17], и наши собственные исследования [11]. Метисированность и дисперсность расселения по территории России послужили главной причиной малого объема собранной выборки кумандинцев (15 чел.), в которую включены только те индивиды, у которых все предки на протяжении трех поколений являлись кумандинцами. Тем не менее, необходимо отметить, что хорошая сопоставимость наших данных о генофонде кумандинцев с данными антропологии и этнографии свидетельствует о том, что исследование этого алтайского народа проведено хоть и на малой, но хорошо собранной выборке.
Рисунок 1. Генетические взаимоотношения между алтайскими этносами (метод Уорда)
Хакасы. Хакасские народы по анализируемым ДНК маркерам оказались более близкими друг другу, чем южные алтайцы – северным (см. табл.), за исключением генетических расстояний между кызыльцами и сагайцами (d = 0.0321). О большей гомогенности хакасов, по сравнению с популяциями Алтая, свидетельствуют и исследования в области антропологии [13, 18]. Наибольшее генетическое сходство выявлено между койбалами и кызыльцами (d = 0.0078) и между койбалами и качинцами (d = 0.0077). Эти хакасские народы характеризуются более выраженной монголоидностью по сравнению с сагайцами, которым присуще некоторое отклонение к западносибирскому (по А.Н. Багашеву) или урало-саянскому (по А.И. Ярхо) антропологическому типу. Кроме того, есть сведения о значительной ассимиляции койбалов качинцами [18].
Рисунок 2. Генетические взаимоотношения между хакасскими этносами (метод Уорда)
Матрица генетических расстояний (см. табл.) и построенная дендрограмма (рис. 2) генетических взаимосвязей между качинцами, койбалами, кызыльцами и сагайцами демонстрируют некоторую обособленность сагайцев, отмечаемую, как уже было указано выше, также и антропологами, в том числе и по краниометрическим данным [3]. Среди хакасских этносов по показателю средней удаленности от всех южносибирских популяций сагайцы явно лидируют (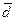 = 0.023), это нашло свое отражение на графике многомерного шкалирования (см. рис. 3). Очевидно, что для понимания причин такого обособленного положения сагайцев необходимо привлечение данных по однородительским ДНК маркерам, которые могут помочь решить вопрос о реальности европеоидного вклада в генофонд хакасов и определить происхождение монголоидного компонента.
= 0.023), это нашло свое отражение на графике многомерного шкалирования (см. рис. 3). Очевидно, что для понимания причин такого обособленного положения сагайцев необходимо привлечение данных по однородительским ДНК маркерам, которые могут помочь решить вопрос о реальности европеоидного вклада в генофонд хакасов и определить происхождение монголоидного компонента.
Шорцы. Две территориальные группы шорцев – горные шорцы Кемеровской области и абаканские шорцы Республики Хакасия – демонстрируют одно из самых небольших генетических расстояний друг относительно друга (d =0.0045), хотя антропологи отмечают заметные различия между ними, причину которого видят в смешении абаканских шорцев с сагайцами и в эффекте основателя [18]. Данные матрицы (см. табл.) и иллюстрации генетических взаимоотношений (см. рис. 3, 4) позволяют говорить о сохраняющемся генетическом сходстве данных территориальных групп и выявляют неожиданную специфику положения шорского генофонда относительно генофонда народов Южной Сибири и Северной Евразии, о чем будет сказано ниже. Величина показателя средней удаленности шорцев от всех изученных южносибирских популяций оказалась выше у шорцев Хакасии ( = 0.018). По этому показателю абаканские шорцы занимают третье место после кумандинцев ( = 0.025) и сагайцев (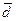 = 0.023).
= 0.023).
ГЕНЕТИЧЕСКИЕ ВЗАИМООТНОШЕНИЯ МЕЖДУ ДЕСЯТЬЮ МАЛЫМИ ЮЖНОСИБИРСКИМИ НАРОДАМИ
Согласно современной антропологической классификации А.Н. Багашева [13], все изученные нами народы (за исключением алтай-кижи, чья таксономическая оценка остается дискуссионной), относятся к разным типам южносибирской (туранской) переходной расы. Северные алтайцы и горные шорцы принадлежат к североалтайскому антропологическому типу, а южные алтайцы, хакасы и абаканские шорцы – к алтае-саянскому.
Одним из основных механизмов генезиса южносибирской расы с точки зрения антропологии явилась метисация – длительное смешение монголоидов и европеоидов [13], при этом состав этнических компонентов, участвовавших в сложении конкретных южносибирских народов, существенно различается. Сопоставление родоплеменной структуры данных народов свидетельствует об определенной общности сеоков (родов), характерной в той или иной степени для всех южносибирских этнических групп, что является отражением истории межэтнических контактов. Отмечалось наличие общих сеоков у койбалов и южных алтайцев [4]; кумандинцев, сагайцев и качинцев; сагайцев и шорцев [14] и так далее. Учитывая сложный механизм этнического сложения народов Южной Сибири, особый интерес представляет исследование генетического сходства малых народов безотносительно к их этнотерриториальным объединениям в алтайцев, хакасов и шорцев (рис. 3). Визуализация генетических связей (см. рис. 3 и 4) выявляет два крупных кластера. Ядром первого являются южные алтайцы, к которым отдельной ветвью присоединяются шорцы, второго – северные алтайцы.
Обращает на себя внимание отсутствие единого кластера хакасских этносов (качинцы, койбалы, кызыльцы, сагайцы), которые на разных уровнях присоединяются к кластерам северных и южных алтайцев. Данные дендрограммы (см. рис. 3) и графика двухмерного шкалирования (см. рис. 4), по всей видимости, отражают сохраняющуюся генетическую гетерогенность хакасов, что согласуется с тезисом об их неоднородности, в пользу которого свидетельствуют и краниометрические характеристики [16].
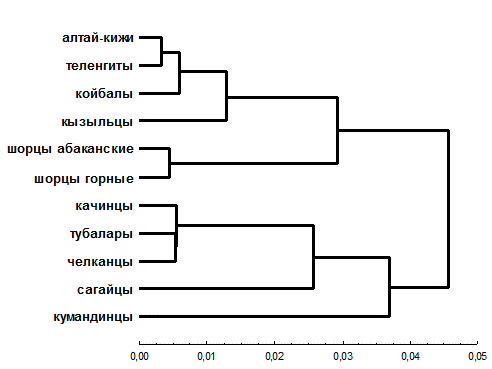
Рисунок 3. Генетические взаимоотношения между десятью южносибирскими народами (метод Уорда)
Сагайцы и качинцы сближаются с северными алтайцами, однако качинцы по результатам шкалирования (см. рис. 4) оказываются все же ближе к койбалам и другим группам алтае-саянского антропологического типа, а их небольшой сдвиг в направлении северных алтайцев может объясняться некоторым ослаблением монголоидных особенностей. Кызыльцы и койбалы – наиболее монголоидные из хакасских народов [18] – объединяются в один кластер с южными алтайцами, для которых также характерна большая выраженность монголоидных черт. Все это ставит под сомнение правомерность объединения этнических хакасских групп в одно этнотерриториальное объединение при молекулярно-генетических исследованиях.


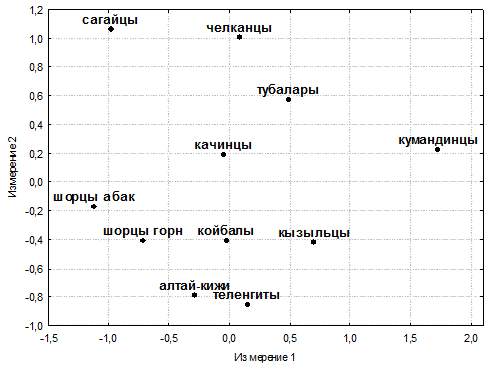
Рисунок 4. Взаимоотношения изученных южносибирских народов
(диаграмма двумерного шкалирования; величина стресса SO = 0.074; коэффициент отчуждения КO = 0.102)
В заключение этой части необходимо отметить, что полученная на основе матрицы генетических расстояний визуализация генетических взаимоотношений десяти южносибирских народов более соответствует таксономии сибирских монголоидов А.Н. Багашева [13], чем Г.Ф. Дебеца [6], который относил шорцев, хакасов, теленгитов и северных алтайцев к одной расе – уральской. Так, и на дендрограмме (см. рис. 3), и на графике двухмерного шкалирования (см. рис. 4) достаточно четко просматриваются два кластера южносибирских этносов, соответствующие, алтае-саянскому и североалтайскому антропологическим типам южносибирской расы. Единственное несоответствие демонстрируют горные шорцы Кемеровской области, которые вместе с абаканскими шорцами Таштыпского района Республики Хакасия оказываются в одном кластере с этносами алтае-саянского типа. Для понимания причин такого положения горных шорцев необходимо дальнейшее изучение генофонда данного народа.
НАРОДЫЮЖНОЙ СИБИРИ В СИСТЕМЕ ГЕНОФОНДА СЕВЕРНОЙ АЗИИ
На следующем шаге анализа нами были рассмотрены генетические взаимоотношения изученных народов Южной Сибири среди различных народов Северной Азии (рис. 5 и 6). К сожалению, из спектра изученных аутосомных ДНК маркеров пришлось исключить один маркер (Alu-инсерция В65), по которому нет информации для популяций бурят, тувинцев и якутов. Кроме того, исключены малочисленные выборки кумандинцев и кызыльцев, а территориальные группы абаканских и горных шорцев объединены в единую группу «шорцы».
На дендрограмме (см. рис. 5) просматриваются несколько кластеров, которые поэтапно объединяются друг с другом. Первые два кластера включают в свой состав этносы, отличающиеся по соотношению монголоидного и европеоидного компонентов. Алтай-кижи, теленгиты, койбалы и якуты – этносы, характеризующиеся выраженной монголоидностью, формируют первый, а тубалары, челканцы, киргизы и качинцы – второй кластер. Вызывает интерес кластеризация качинцев и современных киргизов, возможно, это след вклада в генофонд качинцев енисейских кыргызов, так как по свидетельству этнографической литературы, средневековая элитарная группа кыргызов, сохранившаяся в хакасских степях, в основном составила Тубинский род качинцев [4]. Третий кластер формируют сагайцы и буряты, они присоединяются к первым двум кластерам на одной из последних ветвей.
Из шести народов привлеченных для сравнения только три: казахи, уйгуры и тувинцы образовали отдельный кластер. Буряты, киргизы и якуты объединились, каждый в своем кластере, с южносибирскими народами, что, подтверждает тезис о значительной генетической гетерогенности популяций Южной Сибири.
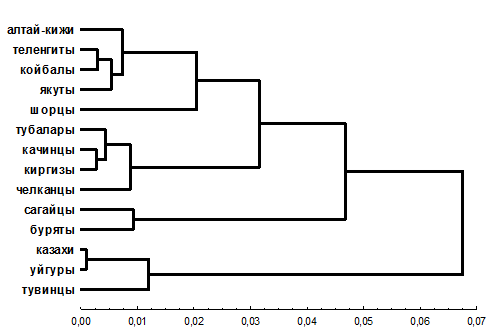
Рисунок 5. Генетические взаимоотношения между шестнадцатью народами Северной Азии (метод Уорда; 8 локусов)
В центре графика двухмерного шкалирования (см. рис. 6) достаточно близко друг относительно друга фокусируется значительная часть проанализированных евразийских народов, за исключением сагайцев, шорцев, челканцев, тувинцев и бурят. По показателю усредненного расстояния или удаленности от всех евразийских популяций самыми дистанцированными оказываются тувинцы (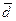 = 0.030), сагайцы ( = 0.028) и буряты ( = 0.024), что, вероятно, является следствием особенностей их формирования на базе разнородных по происхождению генетических компонентов.
= 0.030), сагайцы ( = 0.028) и буряты ( = 0.024), что, вероятно, является следствием особенностей их формирования на базе разнородных по происхождению генетических компонентов.



Рисунок 6. Взаимоотношения изученных южносибирских народов (диаграмма двумерного шкалирования; величина стресса SO = 0.099; коэффициент отчуждения КO = 0.134)
Таким образом, проведенный анализ соотношения генофондов малых народов, входящих в состав алтайцев, хакасов и шорцев, а также оценка их положения в системе генофондов народов Северной Евразии позволяет сделать обобщающий вывод об общности этногенеза и сохраняющейся генетической гетерогенности южносибирских этносов, которая отражается в изменчивости генетических расстояний между малыми народами.
Работа частично поддержана грантом РФФИ 10-04-98002-р_сибирь_а
СПИСОК ЛИТЕРАТУРЫ
- Авакумов А.В. Саяно-Алтайский этногенетический узел (Носители тюхтятской культуры Великого Кыргызского каганата в свете формирования хакасов и киргизов). URL: https://slavica.abakumov.kiev.ua
- Алексеев В.П., Гофман И.И. Антропология азиатской части СССР. – М., 1984. – 208 с.
- Бахолдина В.Ю. Частные системы краниофациальных признаков как инструмент изучения палеоантропологической дифференциации // Вестник антропологии, 2006. – Вып.14. – С. 263-267
- Бутанаев В.Я. Происхождение хакасских родов и фамилий. – Абакан, 1994. – 93 с.
- Генофонд и геногеография народонаселения / Под ред. Ю.Г. Рычкова: Т. 1. Генофонд населения России и сопредельных стран. – СПб: Наука, 2000. – 611с.
- Дебец Г.Ф. Антропологические исследования в Камчатской области (ТИЭ АН СССР. Новая серия). – М., 1951. – Т. 17. – С. 68-119
- Деренко М.В. Молекулярная филогеография коренного населения Северной Евразии по данным об изменчивости митохондриальной ДНК //Автореф. дис…д.б.н., – М, 2009. – 48с.
8. Дерябин В.Е. Биометрическая обработка антропологических данных с применением компьютерных программ. – М.: ВИНИТИ, 2004. – С. 166
9. Лавряшина М.Б., Ульянова М.В., Васинская О.А. и др. Генетический портрет десяти малых народов Южной Сибири. Сообщение I. Полиморфизм аутосомных ДНК маркеров // Медицинская генетика, 2010. – Т.9. – № 3(93). – С.12-17
- Лавряшина М.Б., Ульянова М.В., Балаганская О.А., Балаганский А.Г., Балановская Е.В. Генетический портрет десяти малых народов Южной Сибири. Сообщение II. Гетерозиготность и подразделенность генофонда по данным об аутосомных ДНК маркерах // Медицинская генетика, 2010. – Т.9. – № 9(99). – С. 16-23
- Лавряшина М.Б., Ульянова М.В., Балановская Е.В. Влияние генетико-демографических процессов на структуру сельских популяций коренного населения Южной Сибири: три неперекрывающихся поколения по данным о демографии и распространении фамилий // Медицинская генетика, 2010. – Т.9. – № 4(94). – С.16-24
- Лузина Ф.А. Наследственный полиморфизм и генетические процессы в коренном населении Горного Алтая: Дис...канд. биол. наук. – М., 1987. – 253с.
- Очерки культурогенеза народов Западной Сибири / Отв. ред. А.Н. Багашев. – Т. 4. Расогенез коренного населения. – Томск: Изд-во Том. ун-та, 1998. – 354с.
- Потапов Л.И. Этнический состав и происхождение алтайцев – Л.: Наука, 1969. – 239с.
- Степанов В.А. Этногеномика населения Северной Евразии. – Томск. Изд-во «Печатная мануфактура», 2002. – 244с.
- Тур С.С., Тишкин А.А. К вопросу о происхождении северных алтайцев и шорцев. Интеграция археологических и этнографических исследований: сб. науч. тр. / Под ред. А.Г. Селезнева, С.С. Тихонова, Н.А. Томилова. – Нальчик; Омск: Изд-во ОмГПУ, 2001. – С. 185-188.
- Тур С.С. Современные потомки носителей пазырыкской культуры // Древности Алтая, 2003. – № 10. – С. 141-151
- Тюркские народы Сибири / отв. Ред. Д.А. Функ, Н.А. Томилов. Ин-т этнологии и антропологии им. Н.Н. Миклухо-Маклая РАН; Омский филиал ин-та антропологии и этнографии СО РАН. – М.: Наука, 2006. – 678с.
- Хитринская И.Ю., Степанов В.А., Пузырев В.П. и др. Генетическая дифференциация населения Средней Азии по данным аутосомных маркеров // Генетика, 2003. – Т. 39. – № 10. – С. 1389-1397
- Шерстова Л.И. Этнополитическая история тюрков Южной Сибири. Томск: Изд-во ТПУ, 1999. – С. 290-291
- Ярхо А.И. Алтае-Саянские тюрки. Антропологический очерк. – Абакан: Хакоблнациздат, 1947. – 148с.
- Antunez-de-Mayolo G., Antunez-de-Mayolo A., Antunez-de-Mayolo P. et al. Phylogenetics of worldwide human populations as determined by polymorphic Alu insertions // Electrophoresis, 2002. – Vol. 23. P. – 3346-3356
- Battilana J., Fagundes N.J., Heller A.H., et. al. Alu insertion polymorphisms in Native Americans and related Asian populations // Ann. Hum. Biol., 2006. – Vol. 33. – N 2. – P.142-160
- Nei M. Molecular evolutionary genetics. – Amsterdam, 1975. – 278p.
- Starikovskaya E.V., Sukernik R.I., Derbenina O.A. et. al. Mitochondrial DNA diversity in indigenous population of the southern extent of Siberia and the origins of Native American haplogroups // Ann. Hum. Genet, 2002. – Vol. 69. – P. 67-89