К нуклеозидциклофосфатам относятся нуклеотиды, у которых одна молекула фосфорной кислоты этерифицирует одновременно две гидроксильные группы углеводного остатка. Практически во всех клетках присутствуют два нуклеозидциклофосфата - аденозин-3',5'- циклофосфат (cAMP) и гуанозин-3',5'-циклофосфат (cGMP). Они являются вторичными посредниками (мессенджерами) в передаче гормонального сигнала в клетку.

6. Строение динуклеотидов: ФАД, НАД+, его фосфата НАДФ+. Их участие в окислительно-восстановительных реакциях.
Наиболее важными представителями этой группы соединений являются никотинамидадениндинуклеотид (NAD, или в русской литературе НАД) и его фосфат (NADP, или НАДФ). Эти соединения выполняют важную роль коферментов в осуществлении многих окислительно-восстановительных реакций. В соответствии с этим они могут существовать как в окисленной (НАД+, НАДФ+), так и восстановленной (НАДН, НАДФН) форме.
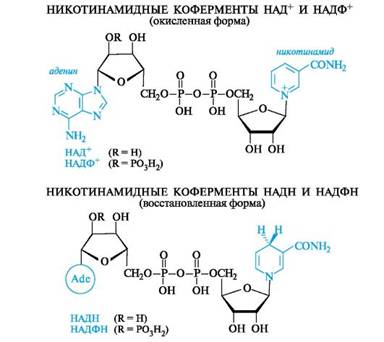
Структурным фрагментом НАД+ и НАДФ+ является никотинамидный остаток в виде пиридиниевого катиона. В составе НАДН и НАДФН этот фрагмент превращается в остаток 1,4-дигидропиридина.
В ходе биологического дегидрирования субстрат теряет два атома водорода, т.е. два протона и два электрона (2Н+, 2е) или протон и гидрид-ион (Н+ и Н-). Кофермент НАД+ обычно рассматривается, как акцептор гидрид-иона Н- (хотя окончательно не установлено, происходит ли перенос атома водорода к этому коферменту одновременно с переносом электрона или эти процессы протекают раздельно).

В результате восстановления путем присоединения гидрид-иона к НАД+ пиридиниевое кольцо переходит в 1,4-дигидропиридиновый фрагмент. Этот процесс обратим.
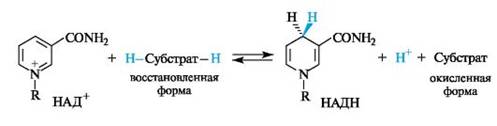
В реакции окисления ароматический пиридиниевый цикл переходит в неароматический 1,4-дигидропиридиновый цикл. В связи с потерей ароматичности возрастает энергия НАДН по сравнению с НАД+. Таким способом НАДН запасает энергию, которая затем расходуется в других биохимических процессах, требующих энергетических затрат.
Типичными примерами биохимических реакций с участием НАД+ служат окисление спиртовых групп в альдегидные (например, превращение этанола в этаналь), а с участием НАДН - восстановление карбонильных групп в спиртовые (превращение пировиноградной кислоты в молочную).
Реакция окисления этанола с участием кофермента НАД+:

В ходе окисления, субстрат теряет два атома водорода, т.е. два протона и два электрона. Кофермент НАД+, приняв два электрона и протон восстанавливается до НАДН при этом нарушается ароматичность. Эта реакция обратима.
При переходе окисленной формы кофермента в восстановленную происходит накопление энергии, выделяемой при окислении субстрата. Накопленная восстановленной формой энергия затем расходуется в других эндэргонических процессах с участием этих коферментов.
ФАД - флавинадениндинуклеотид - кофермент, принимающий участие во многих окислительно-восстановительных биохимических процессах. ФАД существует в двух формах - окисленной и восстановленной, его биохимическая функция, как правило, заключается в переходе между этими формами.
ФАД может быть восстановлен до ФАДH2, при этом он принимает два атома водорода. 
Молекула ФАДH2 является переносчиком энергии, и восстановленный кофермент может быть использован, как субстрат в реакции окислительного фосфорилирования в митохондрии. Молекула ФАДH2 окисляется в ФАД, при этом выделяется энергия, эквивалентная (запасаемая в форме) двум молям ATФ.
Основной источник восстановленного ФАД у эукариот - цикл Кребса и β-окисление липидов. В цикле Кребса ФАД является простетической группой фермента сукцинатдегидрогеназы, которая окисляет сукцинат до фумарата, в β -окислении липидов ФАД является коферментом ацил-CoA дегидрогеназы.
ФАД образуется из рибофлавина, многие оксидоредуктазы, называемые флавопротеинами, для своей работы используют ФАД как простетическую группу в реакциях переноса электронов.
7. Первичная структура нуклеиновых кислот: нуклеотидный состав РНК и ДНК, фосфодиэфирная связь. Гидролиз нуклеиновых кислот.
В полинуклеотидных цепях нуклеотидные звенья связаны через фосфатную группу. Фосфатная группа образует две сложноэфирные связи: с С-3' предыдущего и с С-5' последующего нуклеотидных звеньев (рис. 1). Каркас цепи состоит из чередующихся пентозных и фосфатных остатков, а гетероциклические основания являются «боковыми» группами, присоединенными к пентозным остаткам. Нуклеотид со свободной 5'-ОН группой называют 5'-концевым, а нуклеотид со свободной З'-ОН группой - З'-концевым.

Рис. 1. Общий принцип строения полинуклеотидной цепи
На рисунке 2 приведено строение произвольного участка цепи ДНК, включающего четыре нуклеиновых основания. Легко представить, какое множество сочетаний можно получить путем варьирования последовательности четырех нуклеотидных остатков. Принцип построения цепи РНК такой же, как и у ДНК, с двумя исключениями: пентозным остатком в РНК служит D-рибоза, а в наборе гетероциклических оснований используется не тимин, а урацил.
Первичная структура нуклеиновых кислот определяется последовательностью нуклеотидных звеньев, связанных ковалентными связями в непрерывную цепь полинуклеотида.
Для удобства записи первичной структуры существует несколько способов сокращений. Один из них заключается в использовании ранее приведенных сокращенных названий нуклеозидов. Например, показанный на рис. 2 фрагмент цепи ДНК может быть записан, как d(ApCpGpTp...) или d(A-C-G-T...). Часто букву d опускают, если очевидно, что речь идет о ДНК.

Рис. 2. Первичная структура участка цепи ДНК
Важной характеристикой нуклеиновых кислот служит нуклеотидный состав, т. е. набор и количественное отношение нуклеотидных компонентов. Нуклеотидный состав устанавливают, как правило, путем исследования продуктов гидролитического расщепления нуклеиновых кислот.
ДНК и РНК различаются поведением в условиях щелочного и кислотного гидролиза. ДНК устойчивы к гидролизу в щелочной среде. РНК легко гидролизуются в мягких условиях в щелочной среде до нуклеотидов, которые, в свою очередь, способны в щелочной среде отщеплять остаток фосфорной кислоты с образованием нуклеозидов. Нуклеозиды в кислой среде гидролизуются до гетероциклических оснований и углеводов.

8. Понятие о вторичной структуре ДНК. Комплементарность нуклеиновых оснований. Водородные связи в комплементарных парах нуклеиновых оснований.
Под вторичной структурой понимают пространственную организацию полинуклеотидной цепи. Согласно модели Уотсона-Крика молекула ДНК состоит из двух полинуклеотидных цепей, правозакрученных вокруг общей оси с образованием двойной спирали. Пуриновые и пиримидиновые основания направлены внутрь спирали. Между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи возникают водородные связи. Эти основания составляют комплементарные пары.
Водородные связи образуются между аминогруппой одного основания и карбонильной группой другого -NH...O=C-, а также между амидным и иминным атомами азота -NH...N. Например, как показано ниже, между аденином и тимином образуются две водородные связи, и эти основания составляют комплементарную пару, т. е. аденину в одной цепи будет соответствовать тимин в другой цепи. Другую пару комплементарных оснований составляют гуанин и цитозин, между которыми возникают три водородные связи.

Водородные связи между комплементарными основаниями - один из видов взаимодействий, стабилизирующих двойную спираль. Две цепи ДНК, образующие двойную спираль, не идентичны, но комплементарны между собой. Это означает, что первичная структура, т.е. нуклеотидная последовательность, одной цепи предопределяет первичную структуру второй цепи (рис. 3).

Рис. 3. Комплементарность полинуклеотидных цепей в двойной спирали ДНК
Комплементарность цепей и последовательность звеньев составляют химическую основу важнейшей функции ДНК - хранения и передачи наследственной информации.
В стабилизации молекулы ДНК наряду с водородными связями, действующими поперек спирали, большую роль играют межмолекулярные взаимодействия, направленные вдоль спирали между соседними пространственно сближенными азотистыми основаниями. Поскольку эти взаимодействия направлены вдоль стопки азотистых оснований молекулы ДНК, их называют стэкинг-взаимодействиями. Таким образом, взаимодействия азотистых оснований между собой скрепляют двойную спираль молекулы ДНК и вдоль, и поперек ее оси.
Сильное стэкинг-взаимодействие всегда усиливает водородные связи между основаниями, способствуя уплотнению спирали. Вследствие этого молекулы воды из окружающего раствора связываются в основном с пентозофосфатным остовом ДНК, полярные группы которого находятся на поверхности спирали. При ослаблении стэкинг-взаимодействия молекулы воды, проникая внутрь спирали, конкурентно взаимодействуют с полярными группами оснований, инициируют дестабилизацию и способствуют дальнейшему распаду двойной спирали. Все это свидетельствует о динамичности вторичной структуры ДНК под воздействием компонентов окружающего раствора.

Рис. 4. Вторичная структура молекулы РНК
9. Лекарственные средства на основе модифицированных нуклеиновых оснований (фторурацил, меркаптопурин): структура и механизм действия.
В качестве лекарственных средств в онкологии используют синтетические производные пиримидинового и пуринового рядов, по строению похожие на естественные метаболиты (в данном случае - на нуклеиновые основания), но не полностью им идентичные, т.е. являющиеся антиметаболитами. Например, 5-фторурацил выступает в роли антагониста урацила и тимина, 6-меркаптопурин - аденина. Конкурируя с метаболитами, они нарушают синтез нуклеиновых кислот в организме на разных этапах.
