Тема 13.
Лекция 20. Гормоны
Гормональная регуляция как механизм межклеточной и межорганной координации обмена веществ. Клетки-мишени и клеточные рецепторы гормонов. Гормоны гипоталамуса: либерины и статины.
Гормоны гипофиза. ПОМК как предшественник АКТГ, b-липотропина, эндорфинов. Строение и биологическая роль вазопрессина и окситоцина. Йодсодержащие гормоны, строение и биосинтез. Изменение обмена веществ при гипертиреозе и гипотиреозе. Регуляция фосфорно-кальциевого обмена, участие паратгормона и кальцитонина, активных форм витамина D.
Гормоны поджелудочной железы. Строение, механизм действия инсулина, глюкагона.
Гормоны коры надпочечников: минерало- и глюкокортикоидов. Биосинтез и распад адреналина. Половые гормоны: мужские и женские, влияние на обмен веществ. Гипер- и гипопродукция гормонов.
| Краткое содержание | Конспект | ||
| 1. Гормональная регуляция как механизм межклеточной и межорганной координации обмена веществ. Клетки-мишени и клеточные рецепторы гормонов. Гормоны гипоталамуса: либерины и статины | |||
Учение о гормонах выделено в самостоятельную науку – эндокринологию. Современная эндокринология изучает химическую структуру гормонов, образующихся в железах внутренней секреции, зависимость между структурой и функцией гормонов, молекулярные механизмы действия, а также физиологию и патологию эндокринной системы.
Основные системы регуляции метаболизма и межклеточной коммуникации
1. Центральная и периферическая нервные системы.
2. Эндокринная система через эндокринные железы и гормоны, которые секретируются в кровь и влияют на метаболизм различных клеток-мишеней.
3. Паракринная и аутокринная системы посредством различных соединений, которые секретируются в межклеточное пространство и взаимодействуют с рецепторами либо близлежащих клеток, либо той же клетки (простагландины, гормоны ЖКТ, гистамин и др.).
4. Иммунная система через специфические белки (цитокины, антитела).
3 иерархических уровня системы регуляции обмена веществ и функций организма:
Первый уровень –ЦНС.
Второй уровень –эндокринная система.
Третий уровень –внутриклеточный.
Базальный (нестимулированный) уровень гормонов в крови – в пределах 10-6-10-12 М.
Схема взаимосвязи регуляторных систем организма:
 Существуют эндокринные железы, для которых отсутствуетрегуляция тропными гормонами – паращитовиднаяжелеза, мозговое вещество надпочечников, ренин-альдостероновая система и поджелудочнаяжелеза. Они контролируются нервными влияниями или концентрацией определенных веществ в крови.
Системы регуляции метаболизма:
Существуют эндокринные железы, для которых отсутствуетрегуляция тропными гормонами – паращитовиднаяжелеза, мозговое вещество надпочечников, ренин-альдостероновая система и поджелудочнаяжелеза. Они контролируются нервными влияниями или концентрацией определенных веществ в крови.
Системы регуляции метаболизма:
 Физиологический эффект гормона:
• концентрацией гормона;
• сродством гормонов к белкам-переносчикам;
• количеством рецепторов на поверхности клеток-мишеней;
• типом рецепторов на поверхности клеток-мишеней.
Эволюционно сформировались два основных механизма действия сигнальных молекул в зависимости от локализации рецептора:
1. Мембранный– рецептор расположен на мембране. По данному механизму работают пептидные и белковые гормоны, катехоламины, эйкозаноиды.
2. Цитозольный– рецептор расположен в цитозоле.
Гормоны, связывающиеся с рецепторами на поверхности клетки:
Физиологический эффект гормона:
• концентрацией гормона;
• сродством гормонов к белкам-переносчикам;
• количеством рецепторов на поверхности клеток-мишеней;
• типом рецепторов на поверхности клеток-мишеней.
Эволюционно сформировались два основных механизма действия сигнальных молекул в зависимости от локализации рецептора:
1. Мембранный– рецептор расположен на мембране. По данному механизму работают пептидные и белковые гормоны, катехоламины, эйкозаноиды.
2. Цитозольный– рецептор расположен в цитозоле.
Гормоны, связывающиеся с рецепторами на поверхности клетки:
 Гормоны, связывающиеся с внутриклеточными рецепторами:
Эстрогены, глюкокортикоиды, минералокортикоиды, прогестины, кальцитриол (1,25 [ОН]2-D3), андрогены, тиреоидные гормоны (Т3 и Т4).
Передача гормональных сигналов через внутриклеточные рецепторы:
Гормоны, связывающиеся с внутриклеточными рецепторами:
Эстрогены, глюкокортикоиды, минералокортикоиды, прогестины, кальцитриол (1,25 [ОН]2-D3), андрогены, тиреоидные гормоны (Т3 и Т4).
Передача гормональных сигналов через внутриклеточные рецепторы:
 Гормоны гипоталамуса
К настоящему времени в гипоталамусе открыто 7 стимуляторов (либерины) и 3 ингибитора (статины) секреции гормонов гипофиза, а именно: кортиколиберин, тиролиберин, люлиберин, фоллилиберин, соматолиберин, пролактолиберин, меланолиберин, соматостатин, пролактостатин и меланостатин. По химическому строению все гормоны гипоталамуса являются низкомолекулярными пептидами.
Строение и функции гормонов гипоталамуса:
Гормоны гипоталамуса
К настоящему времени в гипоталамусе открыто 7 стимуляторов (либерины) и 3 ингибитора (статины) секреции гормонов гипофиза, а именно: кортиколиберин, тиролиберин, люлиберин, фоллилиберин, соматолиберин, пролактолиберин, меланолиберин, соматостатин, пролактостатин и меланостатин. По химическому строению все гормоны гипоталамуса являются низкомолекулярными пептидами.
Строение и функции гормонов гипоталамуса:
 Тиролиберин
Пиро-Глу–Гис–Про–NH2
Гонадолиберин
Пиро-Глу–Гис–Трп–Сер–Тир–Гли–Лей–Арг–Про–Гли-NН2
Соматостатин
Тиролиберин
Пиро-Глу–Гис–Про–NH2
Гонадолиберин
Пиро-Глу–Гис–Трп–Сер–Тир–Гли–Лей–Арг–Про–Гли-NН2
Соматостатин
 Соматолиберин
Н-Вал–Гис–Лей–Сер–Ала–Глу–Глн–Лиз–Глу–Ала-ОН
Меланолиберин
Н-Цис–Тир–Иле–Глн–Асн–Цис-ОН
Меланостатин
Пиро-Глу–Гис–Фен–Aрг–Гли–NН2
Местом синтеза гипоталамических гормонов, вероятнее всего, являются нервные окончания – синаптосомы гипоталамуса, поскольку именно там отмечена наибольшая концентрация гормонов и биогенных аминов.
Соматолиберин
Н-Вал–Гис–Лей–Сер–Ала–Глу–Глн–Лиз–Глу–Ала-ОН
Меланолиберин
Н-Цис–Тир–Иле–Глн–Асн–Цис-ОН
Меланостатин
Пиро-Глу–Гис–Фен–Aрг–Гли–NН2
Местом синтеза гипоталамических гормонов, вероятнее всего, являются нервные окончания – синаптосомы гипоталамуса, поскольку именно там отмечена наибольшая концентрация гормонов и биогенных аминов.
| Зарождение науки об эндокринных железах и гормонах относится к 1855 г., когда Т. Аддисон впервые описал бронзовую болезнь, связанную с поражением надпочечников и сопровождающуюся специфической пигментацией кожных покровов. Клод Бернар ввел понятие о железах внутренней секреции, т.е. органах, выделяющих секрет непосредственно в кровь. Позже Ш. Броун-Секар показал, что недостаточность функции желез внутренней секреции вызывает развитие болезней, а экстракты, полученные из этих желез, оказывают хороший лечебный эффект.
Общие свойства гормонов:
- существует 3 класса гормонов (пептидные, стероидные, амины);
- некоторые полипептидные гормоны образуются из неактивных предшественников;
- гормоны действуют в очень низких концентрациях и в большинстве случаев имеют короткое время жизни;
- одни гормоны действуют быстро, другие – медленно;
- гормоны связываются со специфическими рецепторами, расположенными либо на поверхности, либо внутри клетки-мишени;
- действие гормонов может осуществляться через вторичные передатчики (посредники).
Все 75 триллионов клеток, содержащихся в организме человека, служат мишенями одного или нескольких из 50 известных гормонов. Мишенью гормона может быть одна ткань или же несколько тканей. В соответствии с классическим определением ткань-мишень – это такая ткань, в которой гормон вызывает специфическую биохимическую или физиологическую реакцию.
Основные эндокринные системы и их ткани-мишени:
 Обозначить системы регуляции метаболизма:
… - аутокринная система.
… - эндокринная система.
… - паракринная система.
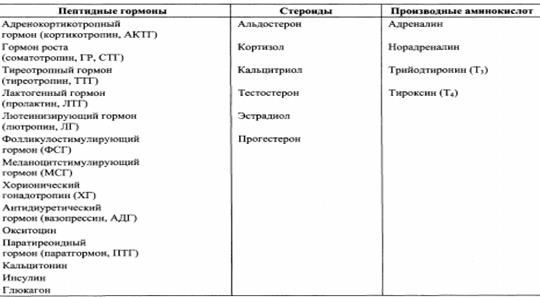
Классификация гормонов организма человека:
Обозначить системы регуляции метаболизма:
… - аутокринная система.
… - эндокринная система.
… - паракринная система.
Классификация гормонов организма человека:
 Пример системы регуляции по типу отрицательной обратной связи Такая система регулирует функцию щитовидной железы, надпочечников, яичников и семенников
Пример системы регуляции по типу отрицательной обратной связи Такая система регулирует функцию щитовидной железы, надпочечников, яичников и семенников
 Основные этапы передачи гормональных сигналов и результат их действия:
Основные этапы передачи гормональных сигналов и результат их действия:

| ||
| 2. Гормоны гипофиза. ПОМК как предшественник АКТГ, b-липотропина, эндорфинов. Строение и биологическая роль вазопрессина и окситоцина. Йодсодержащие гормоны, строение и биосинтез. Изменение обмена веществ при гипертиреозе и гипотиреозе. Регуляция фосфорно-кальциевого обмена, участие паратгормона и кальцитонина, активных форм витамина D. | |||
В зависимости от места синтеза различают гормоны передней, задней и промежуточной долей гипофиза. В передней доле вырабатываются в основном белковые и полипептидные гормоны, называемые тропными гормонами, или тропинами, вследствие их стимулирующего действия на ряд других эндокринных желез.
Строение и биологические функции гормонов передней доли гипофиза:
 Семейство пептидов проопиомеланокортина (ПОМК)
Это семейство состоит из пептидов, действующих либо как гормоны (адренокортикотропин, липотропин, меланоцит-стимулирующий гормон), либо как нейромедиаторы или нейромодуляторы. Проопиомеланокортин (ПОМК) синтезируется в виде молекулы предшественника, состоящей примерно из 285 аминокислотных остатков, и подвергается различному процессингу в разных отделах гипофиза.
Процессинг белка ПОМК в периферических тканях (кишечник, плацента, мужской половой тракт) сходен с таковым в промежуточной доле гипофиза. Существуют три основные группы пептидов семейства ПОМК:
1) АКТГ, из которого могут образовываться меланоцит-стимулирующий гормон (α-МСГ) и кортикотропиноподобный пептид промежуточной доли;
2) β–-липотропин (β–ЛПГ), служащий предшественником α-липотропина, β-МСГ и β-эндорфина и. следовательно, α- и γ-эндорфинов;
3) большой N-концевой пептид, из которого образуетсяγ-МСГ.
Следующее расщепление продуктов ПОМК в передней и промежуточной доле гипофиза происходит на участке между АКТГ и β-ЛПГ, что приводит к отделению N-концевого пептида, включающего АКТГ, от Р-ЛПГ.
АКТГ1-39 затем отделяется от N-концевого пептида, дальнейших расщеплений в передней доле гипофиза практически не происходит. В промежуточной доле АКТГ1-39 расщепляется на α-МСГ (остатки 1-13) и кортикотропиноподобный пептид 18-39); β-липотропин 42-134) превращается в γ-липотропин 42-101) и β-эндорфин 104-134); β-МСГ (84-101) образуется из γ-липотропина.
Перечисленные пептиды претерпевают множество дополнительных модификаций. Большая часть N-концевого пептида и АКТГ1-39 находится в передней доле гипофиза в гликозилированном состоянии, α-МСГ обнаруживается преимущественно в N-ацетилированной и амидированной с С-конца форме; деацетилированный α-МСГ намного менее активен.
β-Эндорфин в промежуточной доле быстро ацетилируется; ацетилированный β-эндорфин в противоположность α-МСГ обладает в 1000 раз меньшей активностью, чем немодифицированная форма. Таким образом, β-эндорфин может находиться в гипофизе в неактивном состоянии. В гипоталамусе молекулы этого пептида не ацетилированы и, по-видимому, присутствуют в активной форме. β-Эндорфин подвергается также укорочению с С-конца с образованием λ- и γ-эндорфинов.
Функции большинства пептидов семейства ПОМК точно не установлены. Постулированные для них эффекты перечислены в табл.
Функции пептидов ПОМК
Семейство пептидов проопиомеланокортина (ПОМК)
Это семейство состоит из пептидов, действующих либо как гормоны (адренокортикотропин, липотропин, меланоцит-стимулирующий гормон), либо как нейромедиаторы или нейромодуляторы. Проопиомеланокортин (ПОМК) синтезируется в виде молекулы предшественника, состоящей примерно из 285 аминокислотных остатков, и подвергается различному процессингу в разных отделах гипофиза.
Процессинг белка ПОМК в периферических тканях (кишечник, плацента, мужской половой тракт) сходен с таковым в промежуточной доле гипофиза. Существуют три основные группы пептидов семейства ПОМК:
1) АКТГ, из которого могут образовываться меланоцит-стимулирующий гормон (α-МСГ) и кортикотропиноподобный пептид промежуточной доли;
2) β–-липотропин (β–ЛПГ), служащий предшественником α-липотропина, β-МСГ и β-эндорфина и. следовательно, α- и γ-эндорфинов;
3) большой N-концевой пептид, из которого образуетсяγ-МСГ.
Следующее расщепление продуктов ПОМК в передней и промежуточной доле гипофиза происходит на участке между АКТГ и β-ЛПГ, что приводит к отделению N-концевого пептида, включающего АКТГ, от Р-ЛПГ.
АКТГ1-39 затем отделяется от N-концевого пептида, дальнейших расщеплений в передней доле гипофиза практически не происходит. В промежуточной доле АКТГ1-39 расщепляется на α-МСГ (остатки 1-13) и кортикотропиноподобный пептид 18-39); β-липотропин 42-134) превращается в γ-липотропин 42-101) и β-эндорфин 104-134); β-МСГ (84-101) образуется из γ-липотропина.
Перечисленные пептиды претерпевают множество дополнительных модификаций. Большая часть N-концевого пептида и АКТГ1-39 находится в передней доле гипофиза в гликозилированном состоянии, α-МСГ обнаруживается преимущественно в N-ацетилированной и амидированной с С-конца форме; деацетилированный α-МСГ намного менее активен.
β-Эндорфин в промежуточной доле быстро ацетилируется; ацетилированный β-эндорфин в противоположность α-МСГ обладает в 1000 раз меньшей активностью, чем немодифицированная форма. Таким образом, β-эндорфин может находиться в гипофизе в неактивном состоянии. В гипоталамусе молекулы этого пептида не ацетилированы и, по-видимому, присутствуют в активной форме. β-Эндорфин подвергается также укорочению с С-конца с образованием λ- и γ-эндорфинов.
Функции большинства пептидов семейства ПОМК точно не установлены. Постулированные для них эффекты перечислены в табл.
Функции пептидов ПОМК
 Гормоны задней доли гипофиза:
вазопрессин(антидиуретический гормон - АДГ), окситоцин.
Оба гормона представляют собой нонапептиды следующего строения:
Гормоны задней доли гипофиза:
вазопрессин(антидиуретический гормон - АДГ), окситоцин.
Оба гормона представляют собой нонапептиды следующего строения:
 Основной биологический эффект окситоцина у млекопитающих связан со стимуляцией сокращения гладких мышц матки при родах и мышечных волокон вокруг альвеол молочных желез, что вызывает секрецию молока.
Вазопрессин стимулирует сокращение гладких мышечных волокон сосудов, оказывая сильное вазопрессорное действие, однако основная роль его в организме сводится к регуляции водного обмена, откуда его второе название антидиуретического гормона. В небольших концентрациях (0,2 нг на 1 кг массы тела) вазопрессин оказывает мощное антидиуретическое действие – стимулирует обратный ток воды через мембраны почечных канальцев. В норме он контролирует осмотическое давление плазмы крови и водный баланс организма человека. При патологии, в частности атрофии задней доли гипофиза, развивается несахарный диабет – заболевание, характеризующееся выделением чрезвычайно больших количеств жидкости с мочой. При этом нарушен обратный процесс всасывания воды в канальцах почек.
Соматотропный гормон (СТГ)
СТГ синтезируется в ацидофильных клетках передней доли гипофиза; концентрация его в гипофизе составляет 5–15 мг на 1 г ткани, что в 1000 раз превышает концентрацию других гормонов гипофиза. К настоящему времени полностью выяснена первичная структура белковой молекулы СТГ человека, быка и овцы. СТГ человека состоит из 191 аминокислоты и содержит две дисульфидные связи; N- и С-концевые аминокислоты представлены фенилаланином.
СТГ обладает широким спектром биологического действия. Он влияет на все клетки организма, определяя интенсивность обмена углеводов, белков, липидов и минеральных веществ. Он усиливает биосинтез белка, ДНК, РНК и гликогена и в то же время способствует мобилизации жиров из депо и распаду высших жирных кислот и глюкозы в тканях. Помимо активации процессов ассимиляции, сопровождающихся увеличением размеров тела, ростом скелета, СТГ координирует и регулирует скорость протекания обменных процессов. Кроме того, СТГ человека и приматов (но не других животных) обладает измеримой лактогенной активностью.
Патологическая биохимия
Гипофункция -при уменьшении выработки и снижении концентрации в крови возникает гипофизарный нанизм (карликовость), частота в России около 1:20000. Проявляется на 2-4 году жизни, критерием служит снижение скорости роста до 4 см в год и менее; при нарушении синтеза у взрослых отмечается снижение мышечной массы и тенденция к гипогликемии; при нарушении рецепции или пострецепторной передачи сигнала происходит пропорциональное, но недостаточное развитие тела. Примером могут служить пигмеи, племена Центральной Африки.
Гиперфункция -у детей возникает гигантизм, так как еще нет зарастания эпифизарных щелей и возможен ускоренный рост кости; у взрослых развивается акромегалия – из-за акрального роста кости происходит увеличение размера носа, стопы, кистей, челюсти.
Адренокортикотропный гормон (АКТГ)
АКТГ вырабатывается базофильными клетками аденогипофиза. АКТГ, помимо основного действия – стимуляции синтеза и секреции гормонов коры надпочечников, обладает жиромобилизующей и меланоцитстимулирующей активностью.
Молекула АКТГ у всех видов животных содержит 39 аминокислотных остатков.
Патологическая биохимия
В результате избыточного образования АКТГ гипофизом или его эктопического образования опухолью развивается синдром Кушинга.Слабое МСГ-подобное действие АКТГ, а также секреция β- или α-МСГ приводят к повышенной пигментации кожи. Возникающие метаболические нарушения обусловлены гиперпродукцией стероидов надпочечников, к ним относятся: 1) отрицательный азотный, калиевый и фосфорный баланс; 2) задержка натрия, которая может привести к повышению артериального давления и отекам; 3) нарушение толерантности к глюкозе или сахарный диабет; 4) повышение содержания жирных кислот в плазме; 5) уменьшение количества эозинофилов и лимфоцитов в крови при увеличении количества полиморфноядерных лейкоцитов. У больных с синдромом Кушинга может наблюдаться атрофия мышц и специфическое перераспределение жира с его отложением на туловище. Отсутствие АКТГ, связанное с опухолью, инфекцией или инфарктом гипофиза, вызывает противоположные сдвиги.
Тиреотропный гормон (ТТГ, тиротропин)
Тиротропин является сложным гликопротеином и содержит, кроме того, по две α- и β-субъединицы, которые в отдельности биологической активностью не обладают: мол. масса его около 30000.
Тиротропин контролирует развитие и функцию щитовидной железы и регулирует биосинтез и секрецию в кровь тиреоидных гормонов.
Предполагают, что действие тиротропина осуществляется, подобно действию других гормонов белковой природы, посредством связывания со специфическими рецепторами плазматических мембран и активирования аденилатциклазной системы.
Липотропные гормоны (ЛТГ, липотропины)
Среди гормонов передней доли гипофиза, структура и функция которых выяснены в последнее десятилетие, следует отметить липотропины, в частности β- и γ-ЛТГ.
К биологическим свойствам β-липотропина относятся жиромобилизующее действие, кортикотропная, меланоцитстимулирующая и гипокальциемическая активность и, кроме того, инсулиноподобный эффект, выражающийся в повышении скорости утилизации глюкозы в тканях.
Среди гормонов передней доли гипофиза, структура и функция которых выяснены в последнее десятилетие, следует отметить липотропины, в частности β- и γ-ЛТГ.
К биологическим свойствам β-липотропина относятся жиромобилизующее действие, кортикотропная, меланоцитстимулирующая и гипокальциемическая активность и, кроме того, инсулиноподобный эффект, выражающийся в повышении скорости утилизации глюкозы в тканях.
Перечисленные биологические свойства обусловлены не β-липотропином, оказавшимся лишенным гормональной активности, а продуктами его распада, образующимися при ограниченном протеолизе. Оказалось, что в ткани мозга и в промежуточной доле гипофиза синтезируются биологически активные пептиды, наделенные опиатоподобным действием.
Структуры некоторых из них приведены ниже:
Н–Тир–Гли–Гли–Фен–Мет–ОН
Метионин-энкефалин
Н–Тир–Гли–Гли–Фен–Лей–ОН
Лейцин-энкефалин
Н–Тир–Гли–Гли–Фен–Мет–Тре–Сер–Глу–Лиз–Сер–Глн–Тре–Про–
Лей–Вал–Тре–Лей–Фен–Лиз–Асн–Ала–Иле–Вал–Лиз–Асн–Ала–Гис–
–Лиз–Лиз–Гли–Глн–ОН
β-Эндорфин
Лактотропный гормон (пролактин, лютеотропный гормон)
Пролактин считается одним из наиболее «древних» гормонов гипофиза. Помимо основного действия (стимуляция развития молочных желез и лактации), пролактин имеет важное биологическое значение – стимулирует рост внутренних органов, секрецию желтого тела (отсюда его второе название «лютеотропный гормон»), оказывает ренотропное, эритропоэтическое и гипергликемическое действие и др. Избыток пролактина, образующийся обычно при наличии опухолей из секретирующих пролактин клеток, приводит к прекращению менструаций (аменорея) и увеличению молочных желез у женщин и к импотенции – у мужчин.
Расшифрована структура пролактина из гипофиза овцы, быка и человека. Это крупный белок, представленный одной полипептидной цепью с тремя дисульфидными связями, состоящий из 199 аминокислотных остатков. Видовые отличия в последовательности аминокислот касаются по существу 2–3 аминокислотных остатков.
В крови женщин уровень пролактина резко повышается перед родами: до 0,2 нг/л против 0,01 нг/л в норме.
Гормоны щитовидной железы
Щитовидная железа состоит из множества особых полостей – фолликулов, заполненных вязким секретом – коллоидом.
В состав коллоида входит особый йодсодержащий гликопротеин с высокой мол. массой – порядка 650000 (5000 аминокислотных остатков). Этот гликопротеин получил название йодтиреоглобулина. Он представляет собой запасную форму тироксина и трийодтиронина – основных гормонов фолликулярной части щитовидной железы.
Структура гормонов щитовидной железы:
Основной биологический эффект окситоцина у млекопитающих связан со стимуляцией сокращения гладких мышц матки при родах и мышечных волокон вокруг альвеол молочных желез, что вызывает секрецию молока.
Вазопрессин стимулирует сокращение гладких мышечных волокон сосудов, оказывая сильное вазопрессорное действие, однако основная роль его в организме сводится к регуляции водного обмена, откуда его второе название антидиуретического гормона. В небольших концентрациях (0,2 нг на 1 кг массы тела) вазопрессин оказывает мощное антидиуретическое действие – стимулирует обратный ток воды через мембраны почечных канальцев. В норме он контролирует осмотическое давление плазмы крови и водный баланс организма человека. При патологии, в частности атрофии задней доли гипофиза, развивается несахарный диабет – заболевание, характеризующееся выделением чрезвычайно больших количеств жидкости с мочой. При этом нарушен обратный процесс всасывания воды в канальцах почек.
Соматотропный гормон (СТГ)
СТГ синтезируется в ацидофильных клетках передней доли гипофиза; концентрация его в гипофизе составляет 5–15 мг на 1 г ткани, что в 1000 раз превышает концентрацию других гормонов гипофиза. К настоящему времени полностью выяснена первичная структура белковой молекулы СТГ человека, быка и овцы. СТГ человека состоит из 191 аминокислоты и содержит две дисульфидные связи; N- и С-концевые аминокислоты представлены фенилаланином.
СТГ обладает широким спектром биологического действия. Он влияет на все клетки организма, определяя интенсивность обмена углеводов, белков, липидов и минеральных веществ. Он усиливает биосинтез белка, ДНК, РНК и гликогена и в то же время способствует мобилизации жиров из депо и распаду высших жирных кислот и глюкозы в тканях. Помимо активации процессов ассимиляции, сопровождающихся увеличением размеров тела, ростом скелета, СТГ координирует и регулирует скорость протекания обменных процессов. Кроме того, СТГ человека и приматов (но не других животных) обладает измеримой лактогенной активностью.
Патологическая биохимия
Гипофункция -при уменьшении выработки и снижении концентрации в крови возникает гипофизарный нанизм (карликовость), частота в России около 1:20000. Проявляется на 2-4 году жизни, критерием служит снижение скорости роста до 4 см в год и менее; при нарушении синтеза у взрослых отмечается снижение мышечной массы и тенденция к гипогликемии; при нарушении рецепции или пострецепторной передачи сигнала происходит пропорциональное, но недостаточное развитие тела. Примером могут служить пигмеи, племена Центральной Африки.
Гиперфункция -у детей возникает гигантизм, так как еще нет зарастания эпифизарных щелей и возможен ускоренный рост кости; у взрослых развивается акромегалия – из-за акрального роста кости происходит увеличение размера носа, стопы, кистей, челюсти.
Адренокортикотропный гормон (АКТГ)
АКТГ вырабатывается базофильными клетками аденогипофиза. АКТГ, помимо основного действия – стимуляции синтеза и секреции гормонов коры надпочечников, обладает жиромобилизующей и меланоцитстимулирующей активностью.
Молекула АКТГ у всех видов животных содержит 39 аминокислотных остатков.
Патологическая биохимия
В результате избыточного образования АКТГ гипофизом или его эктопического образования опухолью развивается синдром Кушинга.Слабое МСГ-подобное действие АКТГ, а также секреция β- или α-МСГ приводят к повышенной пигментации кожи. Возникающие метаболические нарушения обусловлены гиперпродукцией стероидов надпочечников, к ним относятся: 1) отрицательный азотный, калиевый и фосфорный баланс; 2) задержка натрия, которая может привести к повышению артериального давления и отекам; 3) нарушение толерантности к глюкозе или сахарный диабет; 4) повышение содержания жирных кислот в плазме; 5) уменьшение количества эозинофилов и лимфоцитов в крови при увеличении количества полиморфноядерных лейкоцитов. У больных с синдромом Кушинга может наблюдаться атрофия мышц и специфическое перераспределение жира с его отложением на туловище. Отсутствие АКТГ, связанное с опухолью, инфекцией или инфарктом гипофиза, вызывает противоположные сдвиги.
Тиреотропный гормон (ТТГ, тиротропин)
Тиротропин является сложным гликопротеином и содержит, кроме того, по две α- и β-субъединицы, которые в отдельности биологической активностью не обладают: мол. масса его около 30000.
Тиротропин контролирует развитие и функцию щитовидной железы и регулирует биосинтез и секрецию в кровь тиреоидных гормонов.
Предполагают, что действие тиротропина осуществляется, подобно действию других гормонов белковой природы, посредством связывания со специфическими рецепторами плазматических мембран и активирования аденилатциклазной системы.
Липотропные гормоны (ЛТГ, липотропины)
Среди гормонов передней доли гипофиза, структура и функция которых выяснены в последнее десятилетие, следует отметить липотропины, в частности β- и γ-ЛТГ.
К биологическим свойствам β-липотропина относятся жиромобилизующее действие, кортикотропная, меланоцитстимулирующая и гипокальциемическая активность и, кроме того, инсулиноподобный эффект, выражающийся в повышении скорости утилизации глюкозы в тканях.
Среди гормонов передней доли гипофиза, структура и функция которых выяснены в последнее десятилетие, следует отметить липотропины, в частности β- и γ-ЛТГ.
К биологическим свойствам β-липотропина относятся жиромобилизующее действие, кортикотропная, меланоцитстимулирующая и гипокальциемическая активность и, кроме того, инсулиноподобный эффект, выражающийся в повышении скорости утилизации глюкозы в тканях.
Перечисленные биологические свойства обусловлены не β-липотропином, оказавшимся лишенным гормональной активности, а продуктами его распада, образующимися при ограниченном протеолизе. Оказалось, что в ткани мозга и в промежуточной доле гипофиза синтезируются биологически активные пептиды, наделенные опиатоподобным действием.
Структуры некоторых из них приведены ниже:
Н–Тир–Гли–Гли–Фен–Мет–ОН
Метионин-энкефалин
Н–Тир–Гли–Гли–Фен–Лей–ОН
Лейцин-энкефалин
Н–Тир–Гли–Гли–Фен–Мет–Тре–Сер–Глу–Лиз–Сер–Глн–Тре–Про–
Лей–Вал–Тре–Лей–Фен–Лиз–Асн–Ала–Иле–Вал–Лиз–Асн–Ала–Гис–
–Лиз–Лиз–Гли–Глн–ОН
β-Эндорфин
Лактотропный гормон (пролактин, лютеотропный гормон)
Пролактин считается одним из наиболее «древних» гормонов гипофиза. Помимо основного действия (стимуляция развития молочных желез и лактации), пролактин имеет важное биологическое значение – стимулирует рост внутренних органов, секрецию желтого тела (отсюда его второе название «лютеотропный гормон»), оказывает ренотропное, эритропоэтическое и гипергликемическое действие и др. Избыток пролактина, образующийся обычно при наличии опухолей из секретирующих пролактин клеток, приводит к прекращению менструаций (аменорея) и увеличению молочных желез у женщин и к импотенции – у мужчин.
Расшифрована структура пролактина из гипофиза овцы, быка и человека. Это крупный белок, представленный одной полипептидной цепью с тремя дисульфидными связями, состоящий из 199 аминокислотных остатков. Видовые отличия в последовательности аминокислот касаются по существу 2–3 аминокислотных остатков.
В крови женщин уровень пролактина резко повышается перед родами: до 0,2 нг/л против 0,01 нг/л в норме.
Гормоны щитовидной железы
Щитовидная железа состоит из множества особых полостей – фолликулов, заполненных вязким секретом – коллоидом.
В состав коллоида входит особый йодсодержащий гликопротеин с высокой мол. массой – порядка 650000 (5000 аминокислотных остатков). Этот гликопротеин получил название йодтиреоглобулина. Он представляет собой запасную форму тироксина и трийодтиронина – основных гормонов фолликулярной части щитовидной железы.
Структура гормонов щитовидной железы:
 Патологическая биохимия: зоб(увеличение щитовидной железы – недостаток или избыто йодида), гипотиреоз(дефицит свободных Т3 или Т4), гипертиреоз(тиреотоксикоз – избыточное образование тиреоидных гормонов).
Паратиреоидный гормон (ПТГ)
Предшественники и продукты расщепления ПТГ и локализация отдельных этапов расщепления в паращитовидных железах и печени. Цифры в скобках соответствуют числу аминокислот в пре(31) и про(6) фрагментах: →
Патологическая биохимия: недостаток ПТГ приводит к гипопаратиреозу(сниженный уровень ионизированного кальция и повышенный уровень фосфата в сыворотке крови); псевдогипопаратиреоз (органы-мишени резистентны к ПТГ); гиперпаратиреоз (избыточная продукция ПТГ возникает, как правило, вследствие аденомы паратиреоидных желёз); вторичный гиперпаратиреоз (гиперплазия паратиреоидных желёз и гиперсекреция ПТГ).
Кальцитонин (КТ)
Патологическая биохимия: зоб(увеличение щитовидной железы – недостаток или избыто йодида), гипотиреоз(дефицит свободных Т3 или Т4), гипертиреоз(тиреотоксикоз – избыточное образование тиреоидных гормонов).
Паратиреоидный гормон (ПТГ)
Предшественники и продукты расщепления ПТГ и локализация отдельных этапов расщепления в паращитовидных железах и печени. Цифры в скобках соответствуют числу аминокислот в пре(31) и про(6) фрагментах: →
Патологическая биохимия: недостаток ПТГ приводит к гипопаратиреозу(сниженный уровень ионизированного кальция и повышенный уровень фосфата в сыворотке крови); псевдогипопаратиреоз (органы-мишени резистентны к ПТГ); гиперпаратиреоз (избыточная продукция ПТГ возникает, как правило, вследствие аденомы паратиреоидных желёз); вторичный гиперпаратиреоз (гиперплазия паратиреоидных желёз и гиперсекреция ПТГ).
Кальцитонин (КТ)
 Кальцитонин человека
Патологическая биохимия:клинические проявления недостаточности КТ не выявлены. Избыточность КТ наблюдается при медуллярной тиреокарциноме – заболевании, которое может быть спорадическим или семейным.
Биологическое действие кальцитонина прямо противоположно эффекту паратгормона: он вызывает подавление в костной ткани резорбтивных процессов и соответственно гипокальциемию и гипофосфатемию. Таким образом, постоянство уровня кальция в крови человека и животных обеспечивается главным образом паратгормоном, кальцитриолом и кальцитонином.
Биологическое действие гормонов щитовидной железы распространяется на множество физиологических функций организма. В частности, гормоны регулируют скорость основного обмена, рост и дифференцировку тканей, обмен белков, углеводов и липидов, водно-электролитный обмен, деятельность ЦНС, пищеварительного тракта, гемопоэз, функцию сердечнососудистой системы, потребность в витаминах, сопротивляемость организма инфекциям и др. Точкой приложения действия тиреоидных гормонов, как и всех стероидов, считается генетический аппарат.
Специфические рецепторы – белки – обеспечивают транспорт тиреоидных гормонов в ядро и взаимодействие со структурными генами, в результате чего увеличивается синтез ферментов, регулирующих скорость окислительно-восстановительных процессов. Естественно поэтому, что недостаточная функция щитовидной железы (гипофункция) или, наоборот, повышенная секреция гормонов (гиперфункция) вызывает глубокие расстройства физиологического статуса организма.
Гипофункция щитовидной железы в раннем детском возрасте приводит к развитию болезни, известной в литературе как кретинизм. Помимо остановки роста, специфических изменений кожи, волос, мышц, резкого снижения скорости процессов обмена, при кретинизме отмечаются глубокиенарушения психики; специфическое гормональное лечение в этом случае не дает положительных результатов.
Недостаточная функция щитовидной железы в зрелом возрасте сопровождается развитием гипотиреоидного отека, или микседемы (от греч. myxa – слизь, oedemo – отек). Это заболевание чаще встречается у женщин и характеризуется нарушением водно-солевого, основного и жирового обмена.
У больных отмечаются слизистый отек, патологическое ожирение, резкое снижение основного обмена, выпадение волос и зубов, общие мозговые нарушения и психические расстройства. Кожа становится сухой, температура тела снижается; в крови повышено содержание глюкозы. Гипотиреоидизм сравнительно легко поддается лечению препаратами щитовидной железы. Кальцитонин человека
Патологическая биохимия:клинические проявления недостаточности КТ не выявлены. Избыточность КТ наблюдается при медуллярной тиреокарциноме – заболевании, которое может быть спорадическим или семейным.
Биологическое действие кальцитонина прямо противоположно эффекту паратгормона: он вызывает подавление в костной ткани резорбтивных процессов и соответственно гипокальциемию и гипофосфатемию. Таким образом, постоянство уровня кальция в крови человека и животных обеспечивается главным образом паратгормоном, кальцитриолом и кальцитонином.
Биологическое действие гормонов щитовидной железы распространяется на множество физиологических функций организма. В частности, гормоны регулируют скорость основного обмена, рост и дифференцировку тканей, обмен белков, углеводов и липидов, водно-электролитный обмен, деятельность ЦНС, пищеварительного тракта, гемопоэз, функцию сердечнососудистой системы, потребность в витаминах, сопротивляемость организма инфекциям и др. Точкой приложения действия тиреоидных гормонов, как и всех стероидов, считается генетический аппарат.
Специфические рецепторы – белки – обеспечивают транспорт тиреоидных гормонов в ядро и взаимодействие со структурными генами, в результате чего увеличивается синтез ферментов, регулирующих скорость окислительно-восстановительных процессов. Естественно поэтому, что недостаточная функция щитовидной железы (гипофункция) или, наоборот, повышенная секреция гормонов (гиперфункция) вызывает глубокие расстройства физиологического статуса организма.
Гипофункция щитовидной железы в раннем детском возрасте приводит к развитию болезни, известной в литературе как кретинизм. Помимо остановки роста, специфических изменений кожи, волос, мышц, резкого снижения скорости процессов обмена, при кретинизме отмечаются глубокиенарушения психики; специфическое гормональное лечение в этом случае не дает положительных результатов.
Недостаточная функция щитовидной железы в зрелом возрасте сопровождается развитием гипотиреоидного отека, или микседемы (от греч. myxa – слизь, oedemo – отек). Это заболевание чаще встречается у женщин и характеризуется нарушением водно-солевого, основного и жирового обмена.
У больных отмечаются слизистый отек, патологическое ожирение, резкое снижение основного обмена, выпадение волос и зубов, общие мозговые нарушения и психические расстройства. Кожа становится сухой, температура тела снижается; в крови повышено содержание глюкозы. Гипотиреоидизм сравнительно легко поддается лечению препаратами щитовидной железы.
| Подробная схема гипоталамо-гипофизарной системы:
 Продукты расщепления проопиомеланокортина (ПОМК). МСГ – меланоцит-стимулирующий гормон: КПНДГ – кортикотропиноподобный пептид промежуточной доли гипофиза; ЛПГ – липотропин.
Продукты расщепления проопиомеланокортина (ПОМК). МСГ – меланоцит-стимулирующий гормон: КПНДГ – кортикотропиноподобный пептид промежуточной доли гипофиза; ЛПГ – липотропин.
 Патологическая биохимия:
Нарушения секреции или действия АДГ приводят к несахарному диабету, который характеризуется выделением больших объемов разбавленной мочи. Первичный несахарный диабет, связанный с дефицитом АДГ, обычно развивается при повреждении гипоталамо-гипофизарного тракта вследствие перелома основания черепа, опухоли или инфекции: однако он может иметь и наследственную природу. При наследственном нефрогенном несахарном диабете секреция АДГ остается нормальной, но клетки-мишени утрачивают способность реагировать на гормон, вероятно, из-за нарушения его рецепции. Этот наследственный дефект отличается от приобретенного нефрогенного несахарного диабета, который чаще всего возникает при терапевтическом введении лития больным с маниакально-депрессивным психозом. Синдром неадекватной секреции АДГ связан обычно с эктопическим образованием гормона различными опухолями (обычно опухолями легких), но может также наблюдаться и при болезнях мозга, легочных инфекциях или гипотиреозе. Неадекватной такая секреция считается потому, что продукция АДГ происходит с нормальной или повышенной скоростью в условиях гипоосмоляльности, и это вызывает устойчивую и прогрессивную гипонатриемию с выделением гипертонической мочи.
Структура молекулы гормона роста человека. Цифры обозначают положение аминокислотных остатков, начи-
начиная с N-конца.
Патологическая биохимия:
Нарушения секреции или действия АДГ приводят к несахарному диабету, который характеризуется выделением больших объемов разбавленной мочи. Первичный несахарный диабет, связанный с дефицитом АДГ, обычно развивается при повреждении гипоталамо-гипофизарного тракта вследствие перелома основания черепа, опухоли или инфекции: однако он может иметь и наследственную природу. При наследственном нефрогенном несахарном диабете секреция АДГ остается нормальной, но клетки-мишени утрачивают способность реагировать на гормон, вероятно, из-за нарушения его рецепции. Этот наследственный дефект отличается от приобретенного нефрогенного несахарного диабета, который чаще всего возникает при терапевтическом введении лития больным с маниакально-депрессивным психозом. Синдром неадекватной секреции АДГ связан обычно с эктопическим образованием гормона различными опухолями (обычно опухолями легких), но может также наблюдаться и при болезнях мозга, легочных инфекциях или гипотиреозе. Неадекватной такая секреция считается потому, что продукция АДГ происходит с нормальной или повышенной скоростью в условиях гипоосмоляльности, и это вызывает устойчивую и прогрессивную гипонатриемию с выделением гипертонической мочи.
Структура молекулы гормона роста человека. Цифры обозначают положение аминокислотных остатков, начи-
начиная с N-конца.
 Строение АКТГ человека
Н-Сер–Тир–Сер–Мет–Глу–Гис–Фен–Арг–Трп–Гли–Лиз–Про–Вал–Гли–
–Лиз–Лиз–Aрг–Aрг–Про–Вал–Лиз–Вал–Тир–Про–Асп–Ала–Гли–Глу–
–Асп–Глн–Сер–Ала–Глу–Ала–Фен–Про–Лей–Глу–Фен-ОН
Биосинтез гормонов щитовидной железы регулируется тиротропином – гормоном гипоталамуса
Строение АКТГ человека
Н-Сер–Тир–Сер–Мет–Глу–Гис–Фен–Арг–Трп–Гли–Лиз–Про–Вал–Гли–
–Лиз–Лиз–Aрг–Aрг–Про–Вал–Лиз–Вал–Тир–Про–Асп–Ала–Гли–Глу–
–Асп–Глн–Сер–Ала–Глу–Ала–Фен–Про–Лей–Глу–Фен-ОН
Биосинтез гормонов щитовидной железы регулируется тиротропином – гормоном гипоталамуса
 Следует отметить еще одно поражение щитовидной железы – эндемический зоб. Болезнь обычно развивается у лиц, проживающих в горных местностях, где содержание йода в воде и растениях недостаточно. Недостаток йода приводит к компенсаторному увеличению массы ткани щитовидной железы за счет преимущественного разрастания соединительной ткани, однако, этот процесс не сопровождается увеличением секреции тиреоидных гормонов. Болезнь не приводит к серьезным нарушениям функций организма, хотя увеличенная в размерах щитовидная железа создает определенные неудобства. Лечение сводится к обогащению продуктов питания, в частности поваренной соли, неорганическим йодом.
Повышенная функция щитовидной железы (гиперфункция) вызывает развитие гипертиреоза, известного в литературе под названием «зоб диффузный токсический» (болезнь Грейвса, или базедова болезнь). Резкое повышение обмена веществ сопровождается усиленным распадом тканевых белков, что приводит к развитию отрицательного азотистого баланса. Наиболее характерным проявлением болезни считается триада симптомов: резкое увеличение числа сердечных сокращений (тахикардия), пучеглазие (экзофтальм) и зоб, т.е. увеличенная в размерах щитовидная железа; у больных отмечаются общее истощение организма, а также психические расстройства.
Следует отметить еще одно поражение щитовидной железы – эндемический зоб. Болезнь обычно развивается у лиц, проживающих в горных местностях, где содержание йода в воде и растениях недостаточно. Недостаток йода приводит к компенсаторному увеличению массы ткани щитовидной железы за счет преимущественного разрастания соединительной ткани, однако, этот процесс не сопровождается увеличением секреции тиреоидных гормонов. Болезнь не приводит к серьезным нарушениям функций организма, хотя увеличенная в размерах щитовидная железа создает определенные неудобства. Лечение сводится к обогащению продуктов питания, в частности поваренной соли, неорганическим йодом.
Повышенная функция щитовидной железы (гиперфункция) вызывает развитие гипертиреоза, известного в литературе под названием «зоб диффузный токсический» (болезнь Грейвса, или базедова болезнь). Резкое повышение обмена веществ сопровождается усиленным распадом тканевых белков, что приводит к развитию отрицательного азотистого баланса. Наиболее характерным проявлением болезни считается триада симптомов: резкое увеличение числа сердечных сокращений (тахикардия), пучеглазие (экзофтальм) и зоб, т.е. увеличенная в размерах щитовидная железа; у больных отмечаются общее истощение организма, а также психические расстройства.
| ||
| 3. Гормоны поджелудочной железы. Строение, механизм действия инсулина, глюкагона. | |||
Поджелудочная железа относится к железам со смешанной секрецией. Внешнесекреторная функция ее заключается в синтезе ряда ключевых ферментов пищеварения, в частности амилазы, липазы, трипсина, химотрипсина, карбоксипептидазы и др., поступающих в кишечник с соком поджелудочной железы. Внутрисекреторную функцию выполняют, как было установлено в 1902 г. Л.В. Соболевым, панкреатические островки (островки Лангерганса), состоящие из клеток разного типа и вырабатывающие гормоны, как правило, противоположного действия. Так, α- (или А-)клетки продуцируют глюкагон, β- (или В-) клетки синтезируют инсулин, δ-(или D-) клетки вырабатывают соматостатин и F-клетки – малоизученный панкреатический полипептид.
Гормоны поджелудочной железы: инсулин, глюкагон, соматостатин, панкреатический полипептид.
Структура проинсулина и инсулина человека:
 Проинсулин Проинсулин
 Инсулин
Синтезированный из проинсулина инсулин может существовать в нескольких формах, различающихся по биологическим, иммунологическим и физико-химическим свойствам. Различают две формы инсулина: 1) свободную, вступающую во взаимодействие с антителами, полученными к кристаллическому инсулину, и стимулирующую усвоение глюкозы мышечной и жировой тканями; 2) связанную, не реагирующую с антителами и активную только в отношении жировой ткани.
В физиологической регуляции синтеза инсулина доминирующую роль играет концентрация глюкозы в крови. Так, повышение содержания глюкозы в крови вызывает увеличение секреции инсулина в панкреатических островках, а снижение ее содержания, наоборот,– замедление секреции инсулина. Этот феномен контроля по типу обратной связи рассматривается как один из важнейших механизмов регуляции содержания глюкозы в крови.
Механизм действия инсулина окончательно не расшифрован, несмотря на огромное количество фактических данных, свидетельствующих о существовании тесной и прямой зависимости между инсулином и процессами обмена веществ в организме. В соответствии с «унитарной» теорией все эффекты инсулина вызваны его влиянием на обмен глюкозы через фермент гексокиназу. Новые экспериментальные данные свидетельствуют, что усиление и стимуляция инсулином таких процессов, как транспорт ионов и аминокислот, трансляция и синтез белка, экспрессия генов и др., являются независимыми. Это послужило основанием для предположения о множественных механизмах действия инсулина.
Наиболее вероятной в настоящее время представляется мембранная локализация первичного действия почти всех белковых гормонов, включая инсулин. Получены доказательства существования специфического рецептора инсулина на внешней плазматической мембране почти всех клеток организма, а также образования инсулинрецепторного комплекса. Рецептор синтезируется в виде предшественника – полипептида (1382 аминокислотных остатка, мол. масса 190000), который далее расщепляется на α- и β-субъединицы, т.е. на гетеродимер (в формуле α2–β2), связанные дисульфидными связями. Оказалось, что если α-субъединицы (мол. масса 135000) почти целиком располагаются на внешней стороне биомембраны, выполняя функцию связывания инсулина клетки, то β-субъединицы (мол. масса 95000) представляют собой трансмембранный белок, выполняющий функцию преобразования сигнала. Концентрация рецепторов инсулина на поверхности достигает 20000 на клетку, и период их полужизни составляет 7–12 ч.
Патологическая биохимия: сахарный диабет(сахарный диабет I типа – инсулинзависимый, сахарный диабет II типа – инсулин-незавысимый).
Сахарный диабет характеризуется рядом специфических нарушений процессов обмена. Так, у больных развиваются гипергликемия (увеличение уровня глюкозы в крови) и гликозурия (выделение глюкозы с мочой, в которой в норме она отсутствует). К расстройствам обмена относят также усиленный распад гликогена в печени и мышцах, замедление биосинтеза белков и жиров, снижение скорости окисления глюкозы в тканях, развитие отрицательного азотистого баланса, увеличение содержания холестерина и других липидов в крови. При диабете усиливаются мобилизация жиров из депо, синтез углеводов из аминокислот (глюконеогенез) и избыточный синтез кетоновых тел (кетонурия).
Глюкагон
Глюкагон синтезируется в основном в α-клетках панкреатических островков поджелудочной железы, а также в ряде клеток кишечника (см. далее). Он представлен одной линейно расположенной полипептидной цепью, в состав которой входит 29 аминокислотных остатков в следующей последовательности:
Н–Гис–Сер–Глн–Гли–Тре–Фен–Тре–Сер–Асп–Тир–Сер–Лиз–Тир–Лей––Асп–Сер–Aрг–Aрг–Ала–Глн–Асп–Фен–Вал–Глн–Трп–Лей–Мет–Асн––Тре–ОН
Глюкагон образуется из своего предшественника проглюкагона, содержащего на С-конце полипептида дополнительный октапептид (8 остатков), отщепляемый в процессе постсинтетического протеолиза.
По биологическому действию глюкагон, как и адреналин, относятся к гипергликемическим факторам, вызывает увеличение концентрации глюкозы в крови главным образом за счет распада гликогена в печени.
Органами-мишенями для глюкагона являются печень, миокард, жировая ткань, но не скелетные мышцы. Биосинтез и секреция глюкагона контролируются главным образом концентрацией глюкозы по принципу обратной связи. Инсулин
Синтезированный из проинсулина инсулин может существовать в нескольких формах, различающихся по биологическим, иммунологическим и физико-химическим свойствам. Различают две формы инсулина: 1) свободную, вступающую во взаимодействие с антителами, полученными к кристаллическому инсулину, и стимулирующую усвоение глюкозы мышечной и жировой тканями; 2) связанную, не реагирующую с антителами и активную только в отношении жировой ткани.
В физиологической регуляции синтеза инсулина доминирующую роль играет концентрация глюкозы в крови. Так, повышение содержания глюкозы в крови вызывает увеличение секреции инсулина в панкреатических островках, а снижение ее содержания, наоборот,– замедление секреции инсулина. Этот феномен контроля по типу обратной связи рассматривается как один из важнейших механизмов регуляции содержания глюкозы в крови.
Механизм действия инсулина окончательно не расшифрован, несмотря на огромное количество фактических данных, свидетельствующих о существовании тесной и прямой зависимости между инсулином и процессами обмена веществ в организме. В соответствии с «унитарной» теорией все эффекты инсулина вызваны его влиянием на обмен глюкозы через фермент гексокиназу. Новые экспериментальные данные свидетельствуют, что усиление и стимуляция инсулином таких процессов, как транспорт ионов и аминокислот, трансляция и синтез белка, экспрессия генов и др., являются независимыми. Это послужило основанием для предположения о множественных механизмах действия инсулина.
Наиболее вероятной в настоящее время представляется мембранная локализация первичного действия почти всех белковых гормонов, включая инсулин. Получены доказательства существования специфического рецептора инсулина на внешней плазматической мембране почти всех клеток организма, а также образования инсулинрецепторного комплекса. Рецептор синтезируется в виде предшественника – полипептида (1382 аминокислотных остатка, мол. масса 190000), который далее расщепляется на α- и β-субъединицы, т.е. на гетеродимер (в формуле α2–β2), связанные дисульфидными связями. Оказалось, что если α-субъединицы (мол. масса 135000) почти целиком располагаются на внешней стороне биомембраны, выполняя функцию связывания инсулина клетки, то β-субъединицы (мол. масса 95000) представляют собой трансмембранный белок, выполняющий функцию преобразования сигнала. Концентрация рецепторов инсулина на поверхности достигает 20000 на клетку, и период их полужизни составляет 7–12 ч.
Патологическая биохимия: сахарный диабет(сахарный диабет I типа – инсулинзависимый, сахарный диабет II типа – инсулин-незавысимый).
Сахарный диабет характеризуется рядом специфических нарушений процессов обмена. Так, у больных развиваются гипергликемия (увеличение уровня глюкозы в крови) и гликозурия (выделение глюкозы с мочой, в которой в норме она отсутствует). К расстройствам обмена относят также усиленный распад гликогена в печени и мышцах, замедление биосинтеза белков и жиров, снижение скорости окисления глюкозы в тканях, развитие отрицательного азотистого баланса, увеличение содержания холестерина и других липидов в крови. При диабете усиливаются мобилизация жиров из депо, синтез углеводов из аминокислот (глюконеогенез) и избыточный синтез кетоновых тел (кетонурия).
Глюкагон
Глюкагон синтезируется в основном в α-клетках панкреатических островков поджелудочной железы, а также в ряде клеток кишечника (см. далее). Он представлен одной линейно расположенной полипептидной цепью, в состав которой входит 29 аминокислотных остатков в следующей последовательности:
Н–Гис–Сер–Глн–Гли–Тре–Фен–Тре–Сер–Асп–Тир–Сер–Лиз–Тир–Лей––Асп–Сер–Aрг–Aрг–Ала–Глн–Асп–Фен–Вал–Глн–Трп–Лей–Мет–Асн––Тре–ОН
Глюкагон образуется из своего предшественника проглюкагона, содержащего на С-конце полипептида дополнительный октапептид (8 остатков), отщепляемый в процессе постсинтетического протеолиза.
По биологическому действию глюкагон, как и адреналин, относятся к гипергликемическим факторам, вызывает увеличение концентрации глюкозы в крови главным образом за счет распада гликогена в печени.
Органами-мишенями для глюкагона являются печень, миокард, жировая ткань, но не скелетные мышцы. Биосинтез и секреция глюкагона контролируются главным образом концентрацией глюкозы по принципу обратной связи.
|
 Местом синтеза проинсулина считается фракция микросом β-клеток панкреатических островков; превращение неактивного проинсулина в активный инсулин (наиболее существенная часть синтеза) происходит при перемещении проинсулина от рибосом к секреторным гранулам путем частичного протеолиза (отщепление с С-конца полипептидной цепи пептида, содержащего 33 аминокислотных остатка и получившего наименование соединяющего пептида, или С-пептида).
Метаболические сдвиги при инсулиновой недостаточности (сахарный диабет):
Местом синтеза проинсулина считается фракция микросом β-клеток панкреатических островков; превращение неактивного проинсулина в активный инсулин (наиболее существенная часть синтеза) происходит при перемещении проинсулина от рибосом к секреторным гранулам путем частичного протеолиза (отщепление с С-конца полипептидной цепи пептида, содержащего 33 аминокислотных остатка и получившего наименование соединяющего пептида, или С-пептида).
Метаболические сдвиги при инсулиновой недостаточности (сахарный диабет):
 Самым интересным свойством рецептора инсулина, отличным от всех других рецепторов гормонов белковой и пептидной природы, является его способность аутофосфорилирования, т.е. когда рецептор наделен сам протеинкиназной (тирозинкиназной) активностью. При связывании инсулина с α-цепями рецептора происходит активирование тирозинкиназной активности β-цепей путем фосфорилирования их тирозиновых остатков. В свою очередь активная тирозинкиназа β-цепей запускает каскад фосфорилирования–дефосфорилирования протеинкиназ, в частности мембранных или цитозольных серин- или треонинкиназ, т.е. протеинкиназ и белков-мишеней, фосфорилирование в которых осуществляется за счет ОН-групп серина и треонина.
Соответственно имеют место изменения клеточной активности, в частности активация и ингибирование ферментов, транспорт глюкозы, синтез полимерных молекул нуклеиновых кислот и белков и т.д. (Следует указать, что у инсулинрезистентных больных сахарным диабетом синтез инсулина не нарушен, однако организм больных не реагирует ни на свой, ни на инъецированный инсулин. Оказалось, что у части этих больных имеет место мутация в тирозинкиназном домене рецептора и, хотя инсулин связывается нормально с этим мутантным рецептором, дальнейшей передачи сигнала не происходит, так как тирозинкиназа инактивирована. Поэтому лечение больных этой формой диабета инсулином оказывается неэффективным).
В механизме действия глюкагона первичным является связывание со специфическими рецепторами мембраны клеток, образовавшийся глюкагонрецепторный комплекс активирует аденилатциклазу и соответственно образование цАМФ. Последний, являясь универсальным эффектором внутриклеточных ферментов, активирует протеинкиназу, которая в свою очередь фосфорилирует киназу фосфорилазы и гликогенсинтазу. Фосфорилирование первого фермента способствует формированию активной гликогенфосфорилазы и соответственно распаду гликогена с образованием глюкозо-1-фосфата, в то время как фосфорилирование гликогенсинтазы сопровождается переходом ее в неактивную форму и соответственно блокированием синтеза гликогена. Общим итогом действия глюкагона являются ускорение распада гликогена и торможение его синтеза в печени, что приводит к увеличению концентрации глюкозы в крови.
Установлено, что глюкагон способствует образованию глюкозы из промежуточных продуктов обмена белков и жиров. Глюкагон стимулирует образование глюкозы из аминокислот путем индукции синтеза ферментов глюконеогенеза при участии цАМФ. Глюкагон в отличие от адреналина тормозит гликолитический распад глюкозы до молочной кислоты, способствуя тем самым гипергликемии. Он активирует опосредованно через цАМФ липазу тканей, оказывая мощный липолитический эффект. Существуют и различия в физиологическом действии: в отличие от адреналина глюкагон не повышает кровяного давления и не увеличивает частоту сердечных сокращений.
Самым интересным свойством рецептора инсулина, отличным от всех других рецепторов гормонов белковой и пептидной природы, является его способность аутофосфорилирования, т.е. когда рецептор наделен сам протеинкиназной (тирозинкиназной) активностью. При связывании инсулина с α-цепями рецептора происходит активирование тирозинкиназной активности β-цепей путем фосфорилирования их тирозиновых остатков. В свою очередь активная тирозинкиназа β-цепей запускает каскад фосфорилирования–дефосфорилирования протеинкиназ, в частности мембранных или цитозольных серин- или треонинкиназ, т.е. протеинкиназ и белков-мишеней, фосфорилирование в которых осуществляется за счет ОН-групп серина и треонина.
Соответственно имеют место изменения клеточной активности, в частности активация и ингибирование ферментов, транспорт глюкозы, синтез полимерных молекул нуклеиновых кислот и белков и т.д. (Следует указать, что у инсулинрезистентных больных сахарным диабетом синтез инсулина не нарушен, однако организм больных не реагирует ни на свой, ни на инъецированный инсулин. Оказалось, что у части этих больных имеет место мутация в тирозинкиназном домене рецептора и, хотя инсулин связывается нормально с этим мутантным рецептором, дальнейшей передачи сигнала не происходит, так как тирозинкиназа инактивирована. Поэтому лечение больных этой формой диабета инсулином оказывается неэффективным).
В механизме действия глюкагона первичным является связывание со специфическими рецепторами мембраны клеток, образовавшийся глюкагонрецепторный комплекс активирует аденилатциклазу и соответственно образование цАМФ. Последний, являясь универсальным эффектором внутриклеточных ферментов, активирует протеинкиназу, которая в свою очередь фосфорилирует киназу фосфорилазы и гликогенсинтазу. Фосфорилирование первого фермента способствует формированию активной гликогенфосфорилазы и соответственно распаду гликогена с образованием глюкозо-1-фосфата, в то время как фосфорилирование гликогенсинтазы сопровождается переходом ее в неактивную форму и соответственно блокированием синтеза гликогена. Общим итогом действия глюкагона являются ускорение распада гликогена и торможение его синтеза в печени, что приводит к увеличению концентрации глюкозы в крови.
Установлено, что глюкагон способствует образованию глюкозы из промежуточных продуктов обмена белков и жиров. Глюкагон стимулирует образование глюкозы из аминокислот путем индукции синтеза ферментов глюконеогенеза при участии цАМФ. Глюкагон в отличие от адреналина тормозит гликолитический распад глюкозы до молочной кислоты, способствуя тем самым гипергликемии. Он активирует опосредованно через цАМФ липазу тканей, оказывая мощный липолитический эффект. Существуют и различия в физиологическом действии: в отличие от адреналина глюкагон не повышает кровяного давления и не увеличивает частоту сердечных сокращений.
| ||
| 4. Гормоны коры надпочечников: минерало- и глюкокортикоидов. Биосинтез и распад адреналина. Половые гормоны: мужские и женские, влияние на обмен веществ. Гипер- и гипопродукция гормонов | |||
Гормоны коры надпочечников
Биологически активные кортикостероиды объединяют в 3 основные класса:
1. Глюкокортикоиды (С21-стероиды). Основной глюкокортикоид
Поиск по сайту©2015-2024 poisk-ru.ru
Все права принадлежать их авторам. Данный сайт не претендует на авторства, а предоставляет бесплатное использование. Дата создания страницы: 2017-03-31 Нарушение авторских прав и Нарушение персональных данных |
Поиск по сайту: Читайте также: Деталирование сборочного чертежа Когда производственнику особенно важно наличие гибких производственных мощностей? Собственные движения и пространственные скорости звезд |