Агрин существует в нескольких изоформах, которые возникают вследствие альтернативного сплайсинга одного гена. Мотонейроны, мышечные клетки и шванновские клетки экспрессируют агрин, но только у мотонейронов агрин находится в такой изоформе, которая способна вызывать постсинаптическую дифференцировку. Агрин является большой молекулой (heparan sulfate proteoglyсап), домены которой взаимодействуют с ламииином, белками, связывающими гепарин, дистрогликаном, гепарином и интегринами (рис. 5). Способность индуцировать образование постсинаптической специализации в основном зависит от С-концевого домена.
Ведущая роль агрина в формировании нервно-мышечного соединения наиболее очевидна у мышей, у которых при помощи гомологичной рекомбинации выключена экспрессия гена агрина. При таком выключении гена мышечные волокна выглядят нормально и аксон растет в направлении развивающихся мышц, однако нервно-мышечные соединения не образуются.
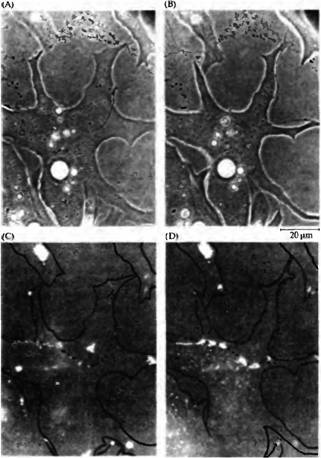
| Рис. 4. Аксоны вызывают агрегацию рецепторов ацетилхолина в областях контакта с мышечными клетками. Фазово контрастные (A. В) и флуоресцентные (С, D) микрофотографии нервно-мышечной культуры клеток Xenopus. Рецепторы ацетилхолина окрашены при помощи родамин бунгаротоксина. (А, С) До и сразу после контакта на миоците имеются спонтанно образованные кластеры рецепторов АХ. (В, D) Через 24 ч спонтанно образовавшиеся области рецепторов АХ исчезли и образовались новые области непосредственно в области аксонального контакта. |
Подобный же фенотип наблюдается у мышей, у которых выключен синтез мышечно-специфического рецептора тирозинкиназы MuSK. Это наводит на мысль, что MuSK формирует часть рецептора к агрину и что вызванное агрином аутофосфорилирование MuSK запускает внутриклеточный сигнальный каскад, который приводит к формированию необходимых компонентов в постсинапсе (рис. 6). Одним из наиболее важных компонентов является рапсин (rapsyn), белок, который, как считается, играет роль в передаче сигналов между рецепторами АХ, MuSK, дистрогликанами и членами Src семейства цитоплазматических рецепторов тирозинкиназ. Таким образом, в нервно-мышечном соединении мутантных мышей с дефицитом рапсина происходит накопление MuSK и выборочная экспрессия гена рецептора АХ в ядре, а также формируются некоторые характерные для синапса свойства, однако не происходит накопления рецепторов АХ.
| Рис. 5. Агрин представляет собой гепаран-сульфат протеогликана большой массы (400-600 кДа). Его домены взаимодействуют с ламинином, гепаран сульфат протеогликанами (HSPGs), гепарином, дистрогликаном, интегрином, гепаринсвязывающими белками и рецепторами агрина, которые вызывают агрегацию рецепторов АХ. (А) Электронная микрофотография агрина после rotatory shadowing. (В) Схематическая диаграмма структурных и связывающихся доменов агрина цыпленка. EG — домен, подобный эпидермальному фактору роста; FS — домен, подобный фолиостатину; LE — домен, подобный ламинину EGF; LG — домен, подобный ламинину G; SEA — последовательность, обнаруженная в белках спермы морского ежа, энтерокиназах и агрине; S/T — домены, богатые серином и/или треонином. Также показаны области связывания. Глобулярные (1, 3-5) и вытянутые (2) области молекулы можно увидеть в части А. | 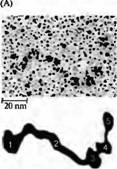
|
Среди белков, которые накапливаются в ответ на изменения в синапсе под действием агрина, находится ARIA, член семейства белков нейрегулинов (neuregulin) и белки рецепторов нейрегулинов erbВ2, erbВЗ и erbВ4. Активация erbВ рецепторов в мышце приводит к экспрессии синаптических субъединиц рецептора АХ.
Гораздо меньше известно о дифференцировке пресинаптической нервной терминали. Эксперименты МакМахана показали, что молекулы, прочно связанные с синаптической базальной мембраной во взрослой мышце, могут вызывать формирование активных зон в регенерирующих аксонах. Отсутствие пресинаптической специализации у мутантных мышей с дефицитом агрина и MuSK может говорить о том, что во время развития пресинаптическая дифференцировка управляется ретроградными сигналами с мышечных клеток в ответ на выделение агрина. Одним из таких ретроградных сигналов, связанных с базальной пластинкой, является ламинин2; он накапливается во время изменений в постсинапсе в ответ на агрин, и у мутантных мышей, у которых имеется дефицит ламинина2, имеются явные аномалии пресинаптической дифференцировки.
| Рис. 6. Взаимодействие агрина с MuSK запускает дифференцировку постсинаптических образований в мышечной клетке, где начинают накапливаться рецепторы АХ, рапсин и дистрогликаны Связывание агрина с MuSK требует корецептора неидентифицированного типа (MASK) и приводит к аутофосфорилированию тирозина MuSK и активации внутриклеточных киназ Src и Fyn. Активированный MuSK захватывает рапсин посредством неидентифицированного трансмембранного белка, RATL. Рапсин, в свою очередь, захватывает дистрогликан и рецепторы АХ, которые фосфорилируются по остаткам тирозина в -субъединице. Через взаимодействие с дистрогликаном происходит накопление большого количества дополнительных синаптических факторов (не показано). | 
|
Выводы
У позвоночных в период эмбриогенеза происходит диффузия протеинов из Шпемановских организационных центров, что приводит к формированию нервной пластинки, края которой загибаются вверх и формируют нервную трубку.
Клетки, расположенные в стенках нервной трубки, быстро делятся. Постмитотические нейроны и клетки-предшественники глии мигрируют в различных направлениях с вентральной поверхности нервной трубки и образуют ЦНС.
Миграция нейронов происходит вдоль радиально расположенных глиальных клеток и клеточных путей, обозначенных различными метками на поверхности клеток и компонентами экстраклеточного матрикса.
Конечное идентифицирование клеток определяется их происхождением и индукционными взаимодействиями с другими клетками.
Гомеотические гены являются управляющими генами, которые контролируют и координируют экспрессию групп других генов и, таким образом, определяют формирование различных частей тела.
Литература:
1. Gilbert, S.F. 2000. Developmental Biology, 6th Ed. Sinauer Associates, Sunderland, MA.
2. Levi-Montalcini, R. 1982. Developmental neuro-biology and the natural history of nerve growth factor. Anny. Rev. Neurosci. 5: 341-362.
3. Lumsden, A, and Krumlauf, R. 1996. Patterning the vertebrate neuroaxis. Science 274: 1109-1115.
4. McAllister, A.K., Katz, L. C., and Lo, D.C. 1999. Neurotrophins and synaptic plasticity. Annu. Rev. Neurosci. 22: 295-318.