Билет 20
Особенности строения и пути проведения информации от болевых и температурных рецепторов
Tерморецепторы
Подразделение:
Ø Два типа: чувствительные к холоду (cold receptors)
и теплу (warm receptors).
Ø Оба типа - свободные нервные окончания.
Проведение импульсов в мозг:
Ø Волокна типа С – немиелинизированные – WARM
Ø Волокна типа Адельта – миелинизированные – COLD
Локализация:
Ø Глубокие слои эпидермиса, дерма, внутр. органы
Ø Рецептивные поля терморецепторов – очень маленькие точки на коже ~ 1 мм в диаметре в коже без волос и до 5 мм в коже с волосами; 3 -4 точки иннервирует 1 нервное окончание.
Физиология:
Ø Чувствительны к скорости изменения температуры, если они быстрые (~5оС в минуту) и, наоборот, медленные изменения (<0,5oC/мин) не вызывают импульсации.
Разряды терморецепторов при разной температуре
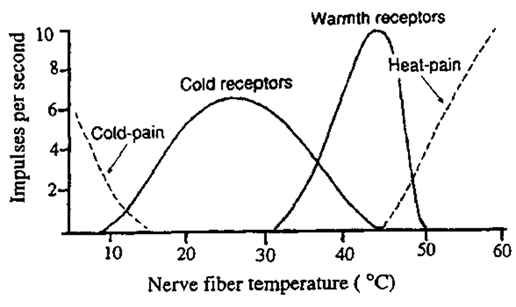
Тонически активны оба типа при 34oC (~2 имп/сек),
но резко увеличивают частоту до 25oC и 45oC соответственно
Тоническая активность температурных рецепторов при неизменной температуре сохраняется в течение некоторого времени, а затем постепенно уменьшается. В таком случае, как и при медленных изменениях действующей на рецепторы температуры, происходит их адаптация, в связи с которой постепенно ослабевают ощущения тепла или холода. Но в ответ на резкие ступенчатые изменения действующей температуры (температурный контраст) возникает залповая физическая активность терморецепторов, которая отражает произошедшие изменения температуры. Наличие адаптации терморецепторов наряду с их способностью реагировать залповой активностью на контрастные изменения действующей температуры проявляются в уменьшении порога тепловой чувствительности при пониженной температуре кожи и в уменьшении порога холодовой чувствительности при повышении ее температуры. Кожные терморецепторы позволяют человеку распознавать нагретые или холодные предметы и окружающий воздух, прикасающиеся к коже. Однако субъективное восприятие, определяемое как «мне холодно» или «мне жарко», возникает в результате суммарной оценки информации, поступившей не только от кожных рецепторов, но также и от терморецепторов спинного мозга, внутренних органов и центральных рецепторов переднего гипоталамуса, где расположен центр терморегуляции. Нервные импульсы от кожных терморецепторов поступают в спинной мозг, в задних рогах которого расположены сенсорные нейроны второго порядка. Их аксоны переходят на противоположную сторону спинного мозга, входя в состав спиноретикулярного и спиноталамического путей. Спиноталамический путь ведет к задним вентролатеральным ядрам таламуса, на нейронах которых конвергируют проводящие пути температурной и тактильной чувствительности. Благодаря такой конвергенции у человека проявляется способность устанавливать, на какую часть поверхности тела подействовал температурный стимул. Спиноретикулярный тракт служит для проведения возбуждения от терморецепторов к ретикулярной формации и принадлежит к неспецифической части сенсорной системы. Ретикулярная формация вместе с неспецифическими ядрами таламуса активирует одновременно многие регионы коры, определяя общую активность мозга и уровень бодрствования. Поэтому повышение притока афферентной информации к ней от терморецепторов, например во время приема холодного душа, оказывает на человека тонизирующее влияние.
Про змей: «Pit organ» (термолокатор)- высокочувствительный терморецептор
Представлен у видов подсемейства Crotalinae (ямкоголовые), семейство гадюковые,Viperidae.
Воспринимает излучения в инфракрасной области 5-30 нм.
Нервный импульс возникает вследствие октрытия – тепло-чувствительных ионных каналов, а не фототрансдукции.
Гипоталамический контроль:
Гипоталамус регулирует температуру тела через взаимодействие с вегетативной, эндокринной и двигательной системами

| Передний гипоталамус (медиальная преоптическая зона) | Задний гипоталамус (зона дорсолатеральнее мамиллярных тел) |
| Центр ТЕПЛООТДАЧИ | Центр ТЕПЛОСОХРАНЕНИЯ |
| Запуск реакций на снижение температуры | Запуск реакций на сохранение тепла |
| Повреждение – гипертермия | Повреждение – гипотермия |
| Электрораздражение – расширение сосудов и подавление дрожи | Электрораздражение – дрожь, вазоконстрикция |
Эндокринные влияния при терморегуляции

Ноцицепция
Болевые рецепторы или ноцицепторы представляют собой свободные нервные окончания, расположенные в коже, слизистых оболочках, мышцах, суставах, надкостнице и во внутренних органах. Чувствительные окончания принадлежат либо безмякотным, либо тонким миелинизиро-ванным волокнам, что определяет скорость проведения сигналов в ЦНС и дает повод к различению ранней боли, короткой и острой, возникающей при проведении импульсов с большей скоростью по миелиновым волокнам, а также поздней, тупой и длительной боли, в случае проведения сигналов по безмякотным волокнам. Ноцицепторы относятся к полимодальным рецепторам, поскольку могут активироваться стимулами разной природы: механическими (удар, порез, укол, щипок), термическими (действие горячих или холодных предметов), химическими (изменение концентрации ионов водорода, действие гистамина, брадикинина и ряда других биологически активных веществ). Порог чувствительности ноцицепторов высок, поэтому лишь достаточно сильные стимулы вызывают возбуждение первичных сенсорных нейронов: например, порог болевой чувствительности для механических стимулов примерно в тысячу раз превышает порог тактильной чувствительности.
Центральные отростки первичных сенсорных нейронов входят в спинной мозг в составе задних корешков и образуют синапсы с нейронами второго порядка, расположенными в задних рогах спинного мозга. Аксоны нейронов второго порядка переходят на противоположную сторону спинного мозга, где образуют спиноталамический и спиноретикулярный тракты. Спиноталамический тракт оканчивается на нейронах нижнего заднелате-рального ядра таламуса, где происходит конвергенция проводящих путей болевой и тактильной чувствительности. Нейроны таламуса образуют проекцию на соматосенсорную кору: этот путь обеспечивает осознаваемое восприятие боли, позволяет определять интенсивность стимула и его локализацию. Волокна спиноретикулярного тракта оканчиваются на нейронах ретикулярной формации, взаимодействующих с медиальными ядрами таламуса. При болевом раздражении нейроны медиальных ядер таламуса оказывают модулирующее влияние на обширные регионы коры и структуры лимбической системы, что приводит к повышению поведенческой активности человека и сопровождается эмоциональными и вегетативными реакциями. Если спиноталамический путь служит для определения сенсорных качеств боли, то спиноретикулярный путь предназначен играть роль сигнала общей тревоги, оказывать на человека общее возбуждающее действие.


Субъективную оценку боли определяет соотношение нейронной активности обоих путей и зависимая от нее активация антиноцицептивных нисходящих путей, способных изменить характер проведения сигналов от ноцицепторов. В сенсорную систему болевой чувствительности встроен эндогенный механизм ее уменьшения путем регуляции порога синаптических переключений в задних рогах спинного мозга («ворота боли»). На передачу возбуждения в этих синапсах влияют нисходящие волокна нейронов серого вещества вокруг водопровода, голубого пятна и некоторых ядер срединного шва. Медиаторы этих нейронов (энкефалин, серотонин, норадреналин) тормозят активность нейронов второго порядка в задних рогах спинного мозга, чем уменьшают проведение афферентных сигналов от ноцицепторов.
Боль, возникающая при раздражении ноцицепторов внутренних органов, называется висцеральной, в отличие от соматической боли, вызванной раздражением болевых рецепторов в коже, мышцах, суставах и костях. Острую висцеральную боль может вызвать сильное растяжение полых органов вследствие закупорки выходных протоков желчного пузыря, мочеточников, в результате непроходимости кишечника, а также спастических сокращений гладких мышц. Плотность ноцицепторов особенно высока в брюшине, при воспалении которой или при ее сильном механическом раздражении возникает острая и интенсивная боль. При воспалительных процессах во внутренних органах выделяются биологически активные вещества, которые служат химическими раздражителями ноцицепторов и причиной болевых ощущений.

Центральные пути болевой чувствительности:


Аналгезирующим (обезболивающим) действием обладают опиатные пептиды (дайнорфин, эндорфины), синтезируемые нейронами гипоталамуса, которые имеют длинные отростки, проникающие в другие отделы головного мозга. Опиатные пептиды присоединяются к специфическим рецепторам нейронов лимбической системы и медиальной области таламуса, их образование увеличивается при некоторых эмоциональных состояниях, стрессе, продолжительных физических нагрузках, у беременных женщин незадолго перед родами, а также в результате психотерапевтического воздействия или акупунктуры. В результате повышенного образования опиатных пептидов активируются антиноцицептивные механизмы и повышается порог болевой чувствительности. Баланс между ощущением боли и ее субъективной оценкой устанавливается с помощью лобных областей мозга, участвующих в процессе восприятия болевых стимулов. При поражении лобных долей (например, в результате травмы или опухоли) порог болевой чувствительности не изменяется и поэтому сенсорный компонент болевого восприятия сохраняется неизменным, однако субъективная эмоциональная оценка боли становится иной: она начинает восприниматься лишь как сенсорное ощущение, а не как страдание.
Физиология глазодвигательной системы
Одна из причин своеобразной организации глазодвигательной системы состоит в том, что механика движений глаз относительно проста и предсказуема, и по этой причине позволяет осуществлять точное предварительное программирование движений. С другой стороны, во многих случаях от этих движений требуется чрезвычайно высокая точность. Кроме того, существуют относительно небольшое количество четко разграниченных классов движений глаз, каждый их которых имеет свои собственные механизмы управления.
Всех позвоночных, обладающих зрением, можно условно разделить на две группы: фовеальные и афовеальные. У фовеальных животных, типичными представителями которых являются приматы, в центре сетчатки имеется специализированная область (у приматов называемая фовеа, или зрительной ямкой), в пределах которой острота зрения значительно повышена по сравнению с остальной частью сетчатки; у афовеальных животных такая специализация сетчатки выражена в значительно меньшей степени. Фовеальное зрение обычно сочетается с фронтальным расположением глаз и объемным бинокулярным зрением, позволяющим значительно точнее воспринимать глубину пространства и определять расстояния до находящихся в нем объектов. Это, в свою очередь, дает возможность более точно производить прыжки – баллистические движения, во время исполнения которых уже практически невозможно исправить ошибки, допущенные при генерации исходной двигательной команды. У афовеальных животных нет необходимости заботиться о точной ориентации зрительной оси глаз, и у них глазодвигательная система выполняет преимущественно функции стабилизации изображения на сетчатке (за счет вестибулоокулярного и оптокинетического рефлексов)
глазодвигательная система позвоночных выполняет следующие функции:
1. Сохранение неподвижного изображения внешнего мира на сетчатке глаза во время движений головы и/или тела животного (у всех позвоночных, обладающих зрением);
2 Помещение проекции выделяющихся объектов зрительного поля в зону сетчатки с высоким разрешением и их прослеживание, а также сканирование целостного изображения внешнего мира (у «фовеальных» животных);
3. Совмещение изображений объектов в двух глазах (у животных с бинокулярным зрением).
В пределах указанных функций можно выделить 6 основных видов движений глаз (видов работы глазодвигательной системы):
1. Вестибулоокулярный рефлекс – стабилизация изображения на сетчатке по сигналам с полукружных каналов;
2. Оптокинетический (оптомоторный) рефлекс – стабилизация изображения на сетчатке по детекции смещения зрительного поля;
3. Саккады – быстрые скачкообразные перемещения глаз;
4. Удерживание взгляда, которое требует тонической работы глазодвигательных мышцы, поскольку упругие силы стремятся вернуть глаз в центральное положение;
5. Плавное прослеживание – перемещение направления взгляда вслед за мелким объектом, движущимся относительно неподвижного фона;
6. Вергенция – сведение и разведение зрительных осей в зависимости от расстояния до объекта.