Трехпетлевая теория Б. Фишера. Основные связи между различными структурами саккадной системы отражает схема, предложенная немецким нейрофизиологом Б. Фишером. Эта схема разработана на основании данных, полученных в следующих экспериментальных условиях: сначала взор устанавливается на фиксационном стимуле, расположенном в центре зрительного поля. Затем появляется второй, периферический стимул, являющийся целью саккады. Задача испытуемого состоит в том, чтобы как можно быстрее перевести взор с центрального стимула на периферический (совершить саккаду). Анализируемым параметром является латентный период саккады – временной интервал между появлением периферического стимула и началом движения.

Опираясь на полученные результаты, Б. Фишер предложил рассматривать систему управления саккадическими движениями глаз как совокупность трех петель, каждой из которых соот-ветствуют определенные морфологические структуры, участвующие в программировании и выполнении саккад.
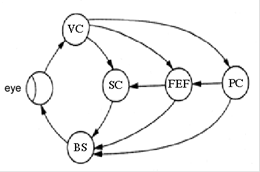
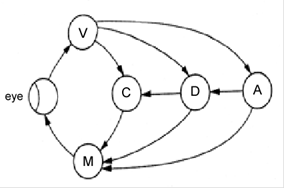
Рисунок. Схема взаимодействия морфологических структур (А) и психофизиологических процессов (Б) в соответствие с трехпетлевой теорией подготовки саккадических движений глаз (Fischer, 1987). VC – зрительная кора, PC – теменная кора, FEF – фронтальное глазодвигательное поле, SC – верхнее дву-холмие, BS – стволовой генератор саккад. V – зрительная ин-формация, A – механизмы внимания, D – принятие решения, C – вычисление координат саккады, M – запуск саккады.
Первая петля замыкается через верхнее двухолмие, отве-чающее за расчет координат саккады (ее направления и ампли-туды), вторая петля проходит через фронтальное глазодвигательное поле, которое связывают с процессами при-нятия решения. И, наконец, третья петля замыкается через те-менную кору, связанную с процессами внимания. Все три петли могут действовать как параллельно, так и последовательно, при этом вторая и третья петли могут тормозить первую до окончания всех процессов, протекающих на более высоком уровне.
Количество петель, задействованных в процессе подготов-ки саккады, может быть различным. От этого будет зависеть длительность латентного периода саккады. Саккады, в подго-товке которых участвует только первая петля, называются экс-пресс-саккадами и имеют латентный период около 90-130 мс. Из них около 55 мс занимает афферентная задержка на сетчат-ке, 25 мс – эфферентная задержка проведения и передачи воз-буждения в нервно-мышечном синапсе, а оставшиеся 10-50 мс – центральное программирование. За это непродолжительное время происходит пересчет ретинотопических координат в про-странственные и активация нейронов верхнего двухолмия.
Увеличение числа структур, задействованных в подготов-ке саккады, приводит к увеличению времени, необходимого для центрального программирования, и удлинению латентного пе-риода. Так, саккады, программируемые с участием фронтально-го глазодвигательного поля (вторая петля), имеют латентный период от 131 до 200 мс и называются коротколатентными. А саккады, в подготовке которых задействованы все три петли, выполняются с латентным периодом более 200 мс – это длин-нолатентные саккады. При этом на центральное программи-рование отводится более 120 мс, в течение которых осуществляются и процессы внимания, и процессы принятия решения, и расчет координат саккады.
Кроме вышеперечисленных, выделяют еще один класс саккад – предугадывающие саккады. Поскольку их латент-ный период слишком мал (менее 90 мс), предугадывающие сак-кады нельзя отнести к движениям, вызванным появлением объекта в поле зрения. Эти саккады связаны с процессами про-гнозирования и представляют собой реакцию, предвосхищаю-щую появление ожидаемого человеком стимула. В программировании предугадывающих саккад участвуют многие структуры саккадной системы. Но, в отличие от саккад других классов, подавляющее большинство процессов, связанных с подготовкой предугадывающих саккад, протекает заранее, в от-сутствие зрительной цели, поэтому такие саккады могут вы-полняться как в первые 90 мс предъявления зрительного стимула, так и до его появления.
Плавное прослеживание
Оптокинетический рефлекс, компенсирует движение всего зрительного поля (или значительной его части), и эта филогенетически древняя система имеется практически у всех позвоночных, обладающих зрением. Однако у фовеальных животных, обладающих специализированной областью сетчатки с высоким разрешением, возникает необходимость стабилизировать изображение лишь на этом небольшом участке зрительного поля, игнорируя его периферию. Эта задача становится самостоятельной в том случае, когда мелкий объект движется относительно фона. При этом улучшение изображения в области фовеа достигается ценой ухудшения восприятия оставшейся большей части зрительного мира (что противоречит оптокинетическому рефлексу), и такой вид движения глаз называют плавным прослеживанием.

Первоначальный захват цели при плавном прослеживании осуществляется саккадой, и это говорит о том, что, видимо, системы плавного прослеживания и саккадная системы тесно связаны друг с другом. В том случае, если движущихся целей две, то вначале направление плавного прослеживания соответствует векторной сумме движения двух объектов, после чего с помощью саккады происходит захват одного из них. Как это ни удивительно, но производить плавное прослеживание в отсутствии реального движущегося стимула можно лишь после длительной тренировки; большинство людей в этой ситуации выполнят лишь саккаду, а не плавное движение глаз. Плавное прослеживание также может быть направлено не только на зрительный стимул, но и на стимулы иной модальности (например, слуховые), хотя его точность при этом намного ниже.
Угловая скорость плавного прослеживания соответствует скорости фиксируемого объекта вплоть до 60-80 °/с, при этом изображение объекта удерживается в пределах 2° от центра фовеа сетчатки. Во время плавного прослеживания нередко возникают небольшие коррекционные саккады, с помощью которых компенсируются возникающие отклонения линии взора от объекта. Если скорость движения объекта достигает 100 °/с и более, то глаз уже систематически отстает от объекта, и по этой причине фрагменты плавного прослеживанию чередуются с большим количеством корректирующих саккад.
Хотя предположительно система плавного прослеживания развилась в эволюции из оптокинетического рефлекса, ее устройство намного сложнее и до сих пор остается недостаточно изученным. Видимо, обработка зрительного сигнала и вычисление моторной команды производится в следующей цепочке: крупноклеточный путь от сетчатки (чувствительный к движению) → латеральное коленчатое тело → первичная зрительная кора → зрительные области, чувствительные к движению (MT, MST) → ядра моста (дорсолатеральное ядро моста) → мозжечок (флоккулонодулярная доля и парафлоккулус) → вестибулярные ядра. Экстрастриарные зрительные области MT и MST специализированы для выделения движущихся объектов из фона; далее сигнал из них попадает в дорсолатеральное ядро моста как непосредственно, так и через заднюю теменную кору и фронтальное глазодвигательное поле. Показано, что дорсолатеральное ядро моста содержит нейроны, кодирующие направление и скорость либо движения мишени, либо ее прослеживания глазами, либо и того и другого одновременно.
Вергенция
Все рассмотренные нами выше движения глаз являются содружественными, т.е. при них оба глаза поворачиваются в одном и том же направлении. В случае вергенции движения двух глаз зеркально симметричны. Предназначение системы вергенции состоит в сведении зрительных осей двух глаз под таким углом, чтобы проекции одного и того же объекта на сетчатках двух глаз совпадали. Если предмет удален на большое расстояние, то это достигается при почти параллельной ориентации зрительных осей, а чем ближе он располагается к голове, тем сильнее должно быть сведение осей (конвергенция), обеспечивающее их пересечение на объекте. Необходимость в вергентных движения возникает лишь у тех животных, которые обладают бинокулярным зрением – среди млекопитающих это, прежде всего, приматы и хищные.
Система вергенции – единственная из систем управления движениями глаз, в которой на два глаза подаются различные двигательные команды. Все остальные системы, включая саккадную и систему плавного прослеживания, выполняют программирование движений глаз таким образом, как если бы оба глаза имели одну общую зрительную ось. Система вергенции обеспечивает "поправку" отдельно для каждого глаза, которая суммируется или вычитается из движения, рассчитанного другими системами.
Вергенция добавляет в систему управления движениями глаз третье измерение – расстояние до объекта. В результате саккаду можно представить уже не в двухмерном пространстве зрительного поля (левее-правее и выше-ниже), а в трехмерном реальном пространстве, включающем расстояние до объекта (ближе-дальше). Рассмотрим в качестве примера, как происходит саккада вправо к новой точке, расположенной ближе к наблюдателю, чем предыдущая точка фиксации. Наблюдаемое при этом движение глаз можно разделить на три фазы. Еще до начала саккады зрительные оси глаз начинают немного сходиться; затем происходит быстрая саккада к новой цели, однако конвергенция зрительных осей при этом даже ускоряется; наконец после остановки саккады замедленное схождение зрительных осей к новому объекту еще продолжается в течение небольшого промежутка времени.
Центр вергенции располагается в ретикулярной формации среднего мозга вблизи от ядра глазодвигательного нерва и организован подобно рассмотренным выше центрам вертикальных и горизонтальных движений глаз. В этой области обнаруживаются пачечные и пачечно-тонические нейроны, которые кодируют само вергентное движение и удерживание глаз в положении определенного сведения зрительных осей. Одни пачечные нейроны разряжаются в связи с конвергенцией, другие – с дивергенцией. Считается, что пачечные нейроны центра вергенции, как и аналогичные нейроны генератора саккад, находятся под постоянным тормозным контролем со стороны клеток-пауз. Во время саккады это торможение снимается, что обусловливает ускорение вергентного движения во время саккады.
Конвергенция зрительных осей представляет лишь один из компонентов рефлекторной триады, возникающей при рассматривании близкорасположенного объекта, и сочетается с увеличением кривизны хрусталика и сужением зрачка. Кстати, и увеличение кривизны хрусталика, и сужение зрачка управляются парасимпатическим ядром Эдингера-Вестфаля, располагающимся в той же самой области среднего мозга вблизи ядра глазодвигательного нерва, а соответствующие преганглионарные парасимпатические волокна направляются к глазу в составе глазодвигательного нерва (III пара).
Основным источником сенсорной информации для осуществления вергентных движений является бинокулярная диспаратность (различия в проекции объемного предмета, видимого двумя глазами под разными углами). Кроме того, независимым источником информации о расстоянии до объекта является степень аккомодации, которая рефлекторно подбирается до получения наиболее четкого изображения (в свою очередь, степень четкости изображения определяется претектальной областью и зрительной корой). Вергенция также может определяться когнитивными факторами, в том числе линейной перспективой и другими признаками глубины пространства. При некоторой тренировке можно научиться производить произвольную дивергенцию зрительных осей независимо от аккомодации. Например, можно научиться рассматривать независимо каждым глазом по одному из двух изображений, составляющих стереоскопическую пару, и добиваться их слияния в трехмерный образ. В полной темноте глаза человека находятся в состоянии конвергенции приблизительно на 3°.