Гипотезы о происхождении вирусов
Существует три основные гипотезы происхождения вирусов[39][40]:
· регрессивная гипотеза;
· гипотеза клеточного происхождения;
· гипотеза коэволюции.
Регрессивная гипотеза
Согласно этой гипотезе, вирусы когда-то были мелкими клетками, паразитирующими в более крупных клетках. С течением времени эти клетки предположительно утратили гены, которые были «лишними» при паразитическом образе жизни. Эта гипотеза основывается на наблюдении, что некоторые бактерии, а именно риккетсии и хламидии, представляют собой клеточные организмы, которые, тем не менее, подобно вирусам могут размножаться только внутри другой клетки. Эту гипотезу также называют гипотезой дегенерации [41][42] или гипотезой редукции [43].
Гипотеза клеточного происхождения
Некоторые вирусы могли появиться из фрагментов ДНК или РНК, которые «высвободились» из генома более крупного организма. Такие фрагменты могут происходить от плазмид (молекул ДНК, способных передаваться от клетки к клетке) или от транспозонов (молекул ДНК, реплицирующихся и перемещающихся с места на место внутри генома)[44]. Транспозоны, которые раньше называли «прыгающими генами», являются примерами мобильных генетических элементов, возможно, от них могли произойти некоторые вирусы. Транспозоны были открыты Барбарой Мак-Клинток в 1950 году в кукурузе[45]. Эту гипотезу также называют гипотезой кочевания [6][46] или гипотезой побега [43].
Гипотеза коэволюции
Эта гипотеза предполагает, что вирусы возникли из сложных комплексов белков и нуклеиновых кислот в то же время, что и первые на Земле живые клетки, и зависят от клеточной жизни вот уже миллиарды лет. Помимо вирусов, существуют и другие неклеточные формы жизни. Например, вироиды — это молекулы РНК, которые не рассматриваются как вирусы, потому что у них нет белковой оболочки. Тем не менее, ряд характеристик сближает их с некоторыми вирусами, а потому их относят к субвирусным частицам[47]. Вироиды являются важными патогенами растений[48]. Они не кодируют собственные белки, однако взаимодействуют с клеткой-хозяином и используют её для осуществления репликации своей РНК[49]. Вирус гепатита D имеет РНК-геном, схожий с геномом вироидов, однако сам не способен синтезировать белок оболочки. Для формирования вирусных частиц он использует белок капсида вируса гепатита B и может размножаться только в клетках, заражённых этим вирусом. Таким образом, вирус гепатита D является дефектным вирусом[50]. Вирофаг Спутник схожим образом зависит от мимивируса, поражающего простейшее Acanthamoeba castellanii [51]. Эти вирусы зависят от присутствия в клетке-хозяине другого вируса и называются вирусами-сателлитами. Подобные вирусы демонстрируют, как может выглядеть промежуточное звено между вирусами и вироидами[52][53].
Каждая из этих гипотез имеет свои слабые места: регрессивная гипотеза не объясняет, почему даже мельчайшие клеточные паразиты никак не походят на вирусы. Гипотеза побега не даёт объяснения появлению капсида и других компонентов вирусной частицы. Гипотеза коэволюции противоречит определению вирусов как неклеточных частиц, зависимых от клеток-хозяев[43].
Тем не менее, в настоящее время многие специалисты признают вирусы древними организмами, появившимися, предположительно, ещё до разделения клеточной жизни на три домена[54]. Это подтверждается тем, что некоторые вирусные белки не обнаруживают гомологии с белками бактерий, архей и эукариот, что свидетельствует о сравнительно давнем обособлении этой группы. В остальном же достоверно объяснить происхождение вирусов на основании трёх закрепившихся классических гипотез не удаётся, что делает необходимыми пересмотр и доработку этих гипотез[54].
Мир РНК
Гипотеза мира РНК[55] и компьютерный анализ последовательностей вирусной ДНК и ДНК хозяина дают лучшее понимание эволюционных взаимоотношений между различными группами вирусов и могут помочь определить предков современных вирусов. До настоящего времени такие исследования пока не прояснили, какая из трёх основных гипотез верна[55]. Однако представляется маловероятным, чтобы все современные вирусы имели общего предка, и, возможно, в прошлом вирусы независимо возникали несколько раз по одному или нескольким механизмам, так как между различными группами вирусов имеются значительные различия в организации генетического материала[56].
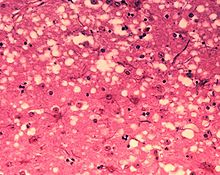
Препарат головного мозга коровы, поражённой коровьим бешенством. В сером веществе образуются микроскопические полости, которые придают ткани вид губки
Прионы
Прионы — это инфекционные белковые молекулы, не содержащие ДНК или РНК[57]. Они вызывают такие заболевания, как почесуха овец[58], губчатая энцефалопатия крупного рогатого скота и хроническая слабость (англ. chronic wasting disease) у оленей. К прионным болезням человека относят куру, болезнь Крейтцфельдта — Якоба и синдром Герстмана — Штраусслера — Шейнкера[59]. Прионы способны стимулировать образование собственных копий. Прионный белок способен существовать в двух изоформах: нормальной (PrPC) и прионной (PrPSc). Прионная форма, взаимодействуя с нормальным белком, способствует его превращению в прионную форму. Хотя прионы фундаментально отличаются от вирусов и вироидов, их открытие даёт больше оснований поверить в то, что вирусы могли произойти от самовоспроизводящихся молекул
Вирусы как форма жизни
Пока вирус находится во внеклеточной среде или в процессе заражения клетки, он существует в виде независимой частицы. Вирусные частицы (вирионы) состоят из двух или трёх компонентов: генетического материала в виде ДНК или РНК (некоторые, например мимивирусы, имеют оба типа молекул); белковой оболочки (капсида), защищающей эти молекулы, и, в некоторых случаях, — дополнительных липидныхоболочек. Наличие капсида отличает вирусы от вирусоподобных инфекционных нуклеиновых кислот — вироидов. В зависимости от того, каким типом нуклеиновой кислоты представлен генетический материал, выделяют ДНК-содержащие вирусы и РНК-содержащие вирусы; на этом принципе основана классификация вирусов по Балтимору. Ранее к вирусам также ошибочно относили прионы, однако впоследствии оказалось, что эти возбудители представляют собой особые инфекционные белки и не содержат нуклеиновых кислот. Форма вирусов варьирует от простой спиральной и икосаэдрической до более сложных структур. Размеры среднего вируса составляют около одной сотой размеров средней бактерии. Большинство вирусов слишком малы, чтобы быть отчётливо различимыми под световым микроскопом.
Вирусы являются облигатными паразитами, так как не способны размножаться вне клетки. Вне клетки вирусные частицы не проявляют признаки живого и ведут себя как частицы биополимеров. От живых паразитарных организмов вирусы отличаются полным отсутствием основного и энергетического обмена и отсутствием сложнейшего элемента живых систем — аппарата трансляции (синтеза белка), степень сложности которого превышает таковую самих вирусов.
Согласно одному из определений вирусы представляют собой форму жизни, согласно другому вирусы являются комплексами органических молекул, взаимодействующими с живыми организмами. Вирусы характеризуют как «организмы на границе живого»[19]. Вирусы похожи на живые организмы в том, что они имеют свой набор генов и эволюционируют путём естественного отбора[61], а также в том, что способны размножаться, создавая собственные копии путём самосборки. Вирусы имеют генетический материал, однако лишены клеточного строения, а именно эту черту обычно рассматривают как фундаментальное свойство живой материи. У вирусов нет собственного обмена веществ, и для синтеза собственных молекул им необходима клетка-хозяин. По этой причине они не способны размножаться вне клетки[62]. В то же время такие бактерии, как риккетсии и хламидии, несмотря на то, что не могут размножаться вне клеток хозяина, считаются живыми организмами[63][64]. Общепризнанные формы жизни размножаются делением клетки, в то время как вирусные частицы самопроизвольно собираются в инфицированной клетке. От роста кристаллов размножение вирусов отличается тем, что вирусы наследуют мутации и находятся под давлением естественного отбора. Самосборка вирусных частиц в клетке даёт дополнительное подтверждение гипотезы, что жизнь могла зародиться в виде самособирающихся органических молекул[3]. Опубликованные в 2013 году данные о том, что некоторые бактериофаги обладают собственной иммунной системой, способной к адаптации[65], являются дополнительным доводом в пользу определения вируса как формы жизни.
Структура
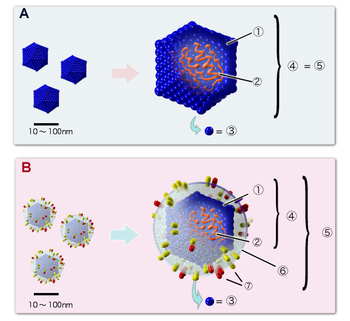 Строение икосаэдрических вирионов: А. Вирус, не имеющий липидной оболочки (например, пикорнавирус). B. Оболочечный вирус (например, герпесвирус). Цифрами обозначены: (1) капсид, (2) геномная нуклеиновая кислота, (3) капсомер, (4) нуклеокапсид, (5) вирион, (6) липидная оболочка, (7) мембранные белки оболочки
Строение икосаэдрических вирионов: А. Вирус, не имеющий липидной оболочки (например, пикорнавирус). B. Оболочечный вирус (например, герпесвирус). Цифрами обозначены: (1) капсид, (2) геномная нуклеиновая кислота, (3) капсомер, (4) нуклеокапсид, (5) вирион, (6) липидная оболочка, (7) мембранные белки оболочки
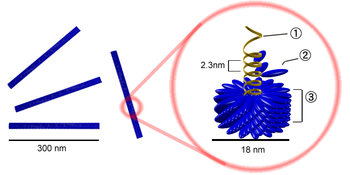 Палочковидный вирион вируса табачной мозаики. Цифрами обозначены: (1) РНК-геном вируса, (2) капсомер, состоящий всего из одного протомера, (3) зрелый участок капсида
Палочковидный вирион вируса табачной мозаики. Цифрами обозначены: (1) РНК-геном вируса, (2) капсомер, состоящий всего из одного протомера, (3) зрелый участок капсида
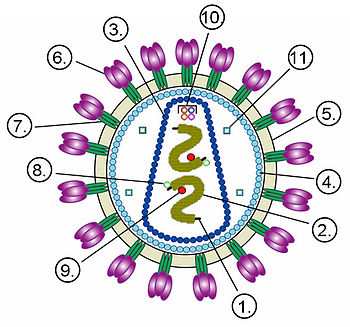 Строение вириона неикосаэдрического оболочечного вируса на примере ВИЧ. Цифрами обозначены: (1) РНК-геном вируса, (2) нуклеокапсид, (3) капсид, (4) белковый матрикс, подстилающий (5) липидную мембрану, (6) gp120 — гликопротеин, с помощью которого происходит связывание вируса с клеточной мембраной, (7) gp41 — гликопротеин. Цифрами 8—11 обозначены белки, входящие в состав вириона и необходимые вирусу на ранних стадиях инфекции: (8) — интеграза, (9) — обратная транскриптаза, (10) — Vif, Vpr, Nef и p7, (11) — протеаза
Строение вириона неикосаэдрического оболочечного вируса на примере ВИЧ. Цифрами обозначены: (1) РНК-геном вируса, (2) нуклеокапсид, (3) капсид, (4) белковый матрикс, подстилающий (5) липидную мембрану, (6) gp120 — гликопротеин, с помощью которого происходит связывание вируса с клеточной мембраной, (7) gp41 — гликопротеин. Цифрами 8—11 обозначены белки, входящие в состав вириона и необходимые вирусу на ранних стадиях инфекции: (8) — интеграза, (9) — обратная транскриптаза, (10) — Vif, Vpr, Nef и p7, (11) — протеаза
|
Вирусы демонстрируют огромное разнообразие форм и размеров. Как правило, вирусы значительно мельче бактерий. Большинство изученных вирусов имеют диаметр в пределах от 20 до 300 нм. Некоторые филовирусы имеют длину до 1400 нм, но их диаметр составляет лишь 80 нм[66]. В 2013 году самым крупным из известных вирусов считался Pandoravirus размерами 1 × 0,5 мкм, однако в 2014 году из многолетней мерзлоты из Сибири был описан Pithovirus, достигающий 1,5 мкм в длину и 0,5 мкм в диаметре. В настоящий момент он считается крупнейшим из известных вирусов[67]. Большинство вирионов невозможно увидеть в световой микроскоп, поэтому используют электронные — как сканирующие, так и просвечивающие[68]. Чтобы вирусы резко выделялись на окружающем фоне, применяют электронноплотные «красители». Они представляют собой растворы солей тяжёлых металлов, таких как вольфрам, которые рассеивают электроны на покрытой ими поверхности. Однако обработка такими веществами ухудшает видимость мелких деталей. В случае негативного контрастирования «окрашивается» только фон[69].
Зрелая вирусная частица, известная как вирион, состоит из нуклеиновой кислоты, покрытой защитной белковой оболочкой — капсидом. Капсид складывается из одинаковых белковых субъединиц, называемых капсомерами [70]. Вирусы могут также иметь липидную оболочку поверх капсида (суперкапсид), образованную из мембраны клетки-хозяина. Капсид состоит из белков, кодируемых вирусным геномом, а его форма лежит в основе классификации вирусов по морфологическому признаку[71][72]. Сложноорганизованные вирусы, кроме того, кодируют специальные белки, помогающие в сборке капсида. Комплексы белков и нуклеиновых кислот известны как нуклеопротеины, а комплекс белков вирусного капсида с вирусной нуклеиновой кислотой называется нуклеокапсидом. Форму капсида и вириона в целом можно механически (физически) исследовать при помощи сканирующего атомно-силового микроскопа[73][74].
Капсид
Классифицируют четыре морфологических типа капсидов вирусов: спиральный, икосаэдрический, продолговатый и комплексный.
Спиральный
Эти капсиды состоят из одного типа капсомеров, уложенных по спирали вокруг центральной оси. В центре этой структуры может находиться центральная полость или канал. Такая организация капсомеров приводит к формированию палочковидных и нитевидных вирионов: они могут быть короткими и очень плотными или длинными и очень гибкими. Генетический материал, как правило, представлен одноцепочечной РНК (в некоторых случаях одноцепочечной ДНК) и удерживается в белковой спирали ионными взаимодействиями между отрицательными зарядами на нуклеиновых кислотах и положительными зарядами на белках. В целом, длина спирального капсида зависит от длины окружённой им нуклеиновой кислоты, а диаметр определяется размером и расположением капсомеров. Примером спирального вируса может служить вирус табачной мозаики[75].
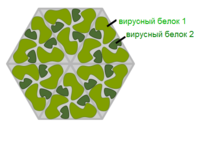
Вирус с капсомером типа гексон. Эта иллюстрация показывает, что вирусный капсид может быть выстроен из множественных копий всего лишь двух белков
Икосаэдрический
Большинство вирусов животных имеют икосаэдрическую или почти шарообразную форму с икосаэдрической симметрией. Правильный икосаэдр является оптимальной формой для закрытого капсида, сложенного из одинаковых субъединиц. Минимальное необходимое число одинаковых капсомеров — 12, каждый капсомер состоит из пяти идентичных субъединиц. Многие вирусы, такие как ротавирус, имеют более двенадцати капсомеров и выглядят круглыми, но сохраняют икосаэдрическую симметрию. Капсомеры, находящиеся в вершинах, окружены пятью другими капсомерами и называются пентонами. Капсомеры треугольных граней имеют 6 соседей-капсомеров и называются гексонами [76]. Гексоны, по существу, являются плоскими, а пентоны, образующие 12 вершин, — изогнутыми. Один и тот же белок может выступать субъединицей и пентомеров, и гексамеров, или же они могут состоять из различных белков.
Продолговатый
Продолговатыми называют икосаэдрические капсиды, вытянутые вдоль оси симметрии пятого порядка. Такая форма характерна для головок бактериофагов[77].
Комплексный
Форма этих капсидов ни чисто спиральная, ни чисто икосаэдрическая. Они могут нести дополнительные наружные структуры, такие как белковые хвосты или сложные наружные стенки. Некоторые бактериофаги, такие как фаг Т4, имеют комплексный капсид, состоящий из икосаэдрической головки, соединённой со спиральным хвостом, который может иметь шестигранное основание с отходящими от него хвостовыми белковыми нитями. Этот хвост действует наподобие молекулярного шприца, прикрепляясь к клетке-хозяину и затем впрыскивая в неё генетический материал вируса[78].
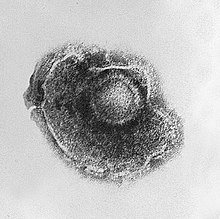
Вирус герпеса, окружённый липидной оболочкой (суперкапсидом)
Оболочка
Некоторые вирусы окружают себя дополнительной оболочкой из модифицированной клеточной мембраны (плазматической или внутренней, такой как ядерная мембрана или мембрана эндоплазматического ретикулума). Этот дополнительный билипидный слой называется суперкапсидом. Липидная оболочка вируса испещрена белками, кодируемыми вирусными геномом и геномом хозяина; сама же мембрана, а также любые её углеводные компоненты происходят целиком из клетки-хозяина. Таким образом формируют свою оболочку вирус гриппа и ВИЧ. Инфекционность большинства вирусов, имеющих оболочку, зависит именно от этой оболочки[79].
Поксвирусы представляют собой крупные сложноорганизованные вирусы с необычной морфологией. Генетический материал вируса связан с белками в центральной дисковидной структуре, известной как нуклеоид. Нуклеоид окружён мембраной и двумя боковыми телами неизвестной функции. Вирус имеет наружную оболочку с большим количеством белков на её поверхности. Весь вирион слегка плейоморфен (то есть способен изменять форму и размер в зависимости от условий) и может принимать форму от овальной до блоковидной[80]. Мимивирус является одним из крупнейших описанных вирусов и имеет икосаэдрический капсид диаметром 400—500 нм. Белковые филаменты, отходящие от поверхности вириона, достигают 100 нм в длину[81][82]. В 2011 году исследователи обнаружили ещё более крупный вирус на океаническом дне вблизи побережья Чили. Вирус, которому было дано временное название Megavirus chilensis, можно увидеть даже в обычный оптический микроскоп[83].
Геном
| Генетическое разнообразие у вирусов | |
| Свойства | Параметры |
| Нуклеиновая кислота | · ДНК · РНК · И ДНК, и РНК (в различных стадиях жизненного цикла) |
| Форма | · Линейная · Кольцевая · Сегментированная |
| Количество цепей | · Одноцепочечные · Двуцепочечные · Двуцепочечные с одноцепочечными фрагментами |
| Полярность | · Положительная полярность (+) · Отрицательная полярность (−) · Двойная полярность (+/−) |
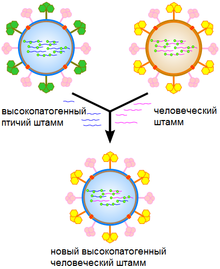
Реассортимент может стать причиной появления нового высокопатогенного штамма человеческого гриппа
Вирусы демонстрируют огромное количество вариантов организации генома; в этом смысле они более разнообразны, чем растения, животные, археи и бактерии. Существуют миллионы различных типов вирусов[8], но только примерно 5000 из них были детально описаны[7]. Генетический материал вируса может быть представлен либо ДНК, либо РНК, соответственно, вирусы подразделяют на ДНК-содержащие и РНК-содержащие. Подавляющее большинство вирусов являются РНК-содержащими. Вирусы растений чаще всего содержат одноцепочечную РНК, а бактериофаги, как правило, обладают двухцепочечными ДНК[84].
Вирусный геном может быть кольцевым, как у полиомавирусов, или линейным, как у аденовирусов. Форма генома не зависит от типа нуклеиновой кислоты. У многих РНК-содержащих вирусов и некоторых ДНК-содержащих вирусов геном часто представлен несколькими молекулами (частями), в связи с чем он называется сегментированным. У РНК-содержащих вирусов каждый сегмент часто кодирует только один белок, и обычно эти сегменты упаковываются в один капсид. Однако присутствие всех сегментов не всегда обязательно для инфекционности вируса, как это демонстрируют вирус мозаики костра[en] и некоторые другие вирусы растений[66].
Вирусные геномы независимо от типа нуклеиновый кислоты практически всегда бывают либо одноцепочечным, либо двухцепочечным. Двухцепочечный геном включает пару комплементарных цепей нуклеиновой кислоты, а одноцепочечный — только одну цепь. Геном вирусов некоторых семейств (например, Hepadnaviridae) частично одноцепочечный и частично двуцепочечный[84].
Для большинства РНК-содержащих вирусов и некоторых вирусов с одноцепочечной ДНК определяют полярность нуклеиновой кислоты в зависимости от того, комплементарна ли она вирусной мРНК. Молекула РНК с положительной полярностью (плюс-цепь) имеет ту же последовательность нуклеотидов, что и мРНК, поэтому, по крайней мере, какая-то её часть может незамедлительно начать транслироваться клеткой-хозяином. РНК с отрицательной полярностью (минус-цепь) комплементарна мРНК, поэтому до начала трансляции на ней должна быть синтезирована положительная РНК при помощи фермента РНК-зависимой-РНК-полимеразы. Названия цепей ДНК для вирусов, содержащих одноцепочечную ДНК, сходны с таковыми для РНК: кодирующая цепь комплементарна мРНК (-), а некодирующая является её копией (+)[84]. Однако геномы нескольких типов ДНК- и РНК-содержащих вирусов представлены молекулами, имеющими различную полярность, то есть транскрипции может подвергаться любая цепь. Таковы, например, геминивирусы — вирусы растений, содержащие одноцепочечную ДНК, — и аренавирусы — вирусы животных с одноцепочечной РНК[85].
Размер генома широко варьирует у различных видов. Самым маленьким одноцепочечным ДНК-геномом обладает цирковирус из семейства Circoviridae: его геном кодирует лишь два белка и содержит всего 2000 нуклеотидов. Один из самых крупных геномов обнаружен у мимивируса: он содержит свыше 1,2 млн пар оснований и кодирует более тысячи белков[86]. Как правило, РНК-содержащие вирусы имеют меньший геном, чем ДНК-содержащие — размер их генома ограничен из-за большей вероятности ошибок во время репликации[38]. При большем размере генома ошибки, произошедшие во время его репликации, сделали бы вирус нежизнеспособным или неконкурентоспособным. Чтобы преодолеть это ограничение, РНК-вирусы часто имеют сегментированный геном — это уменьшает вероятность того, что ошибка в одном из сегментов окажется фатальной для всего генома. Напротив, ДНК-содержащие вирусы обычно имеют более крупные геномы благодаря большей точности их репликативных ферментов[87]. Однако вирусы, содержащие одноцепочечные ДНК, являются исключением из этого правила — скорость накопления мутаций в их геномах приближается к таковой для вирусов, содержащих одноцепочечные РНК[88].
Генетические изменения происходят у вирусов по разным механизмам. В их число входят случайные замены отдельных оснований в РНК или ДНК. В большинстве случаев эти точечные мутации являются «молчащими» — они не изменяют структуру белков, кодируемых мутантными генами, но иногда в результате таких изменений вирус может приобрести эволюционные преимущества, такие как устойчивость к противовирусным препаратам[89]. Дрейф антигенов имеет место тогда, когда в геноме вируса происходят масштабные изменения. Это может быть результатом рекомбинации или реассортимента. Когда это случается с вирусом гриппа, результатом может стать пандемия[90]. РНК-вирусы часто существуют как квазивиды или смесь вирусов одного вида, но с чуть различающимися нуклеотидными последовательностями генома. Такие квазивиды являются главной мишенью для естественного отбора[91].
Сегментированный геном даёт эволюционные преимущества: различные штаммы вируса с сегментированным геномом могут обмениваться генами и производить потомков с уникальными характеристиками. Это явление называется реассортиментом[92].
Генетическая рекомбинация — это процесс внесения разрыва в молекулу нуклеиновой кислоты с последующим «сшиванием» её с другими молекулами нуклеиновой кислоты. Рекомбинация может происходить между геномами двух вирусов, когда они заражают клетку одновременно. Исследования эволюции вирусов показали, что у изученных видов рекомбинация широко распространена[93]. Рекомбинация характерна как для РНК-, так и для ДНК-содержащих вирусов[94][95].
Жизненный цикл
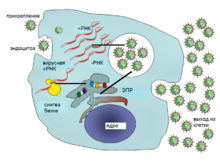
Типичный жизненный цикл вируса на примере вируса гепатита C
Вирусы не размножаются клеточным делением, поскольку не имеют клеточного строения. Вместо этого они используют ресурсы клетки-хозяина для образования множественных копий самих себя, и их сборка происходит внутри клетки.
Условно жизненный цикл вируса можно разбить на несколько взаимоперекрывающихся этапов (обычно выделяют 6 этапов[96]):
· Прикрепление представляет собой образование специфичной связи между белками вирусного капсида и рецепторами на поверхности клетки-хозяина. Это специфичное связывание определяет круг хозяев вируса. Например, ВИЧ поражает только определённый тип человеческих лейкоцитов. Это связано с тем, что оболочечный гликопротеин вируса gp120 специфично связывается с молекулой CD4 — хемокиновымрецептором, который обычно встречается на поверхности CD4+ T-лимфоцитов. Этот механизм обеспечивает инфицирование вирусом только тех клеток, которые способны осуществить его репликацию. Связывание с рецептором может вызвать конформационные изменения белка оболочки (или белка капсида в случае безоболочечного вируса), что в свою очередь служит сигналом к слиянию вирусной и клеточной мембран и проникновению вируса в клетку.
· Проникновение в клетку. На следующем этапе вирусу необходимо доставить внутрь клетки свой генетический материал. Некоторые вирусы также переносят внутрь клетки собственные белки, необходимые для её реализации (особенно это характерно для вирусов, содержащих негативные РНК). Различные вирусы для проникновения в клетку используют разные стратегии: например, пикорнавирусы впрыскивают свою РНК через плазматическую мембрану, а вирионы ортомиксовирусов захватываются клеткой в ходе эндоцитоза и попадают в кислую среду лизосом, где происходит депротеинизация вирусной частицы, после чего РНК в комплексе с вирусными белками преодолевает лизосомальную мембрану и попадает в цитоплазму. Вирусы также различают по тому, где происходит их репликация: часть вирусов (например, те же пикорнавирусы) размножается в цитоплазме клетки, а часть (например, ортомиксовирусы) в её ядре. Процесс инфицирования вирусами клеток грибов и растений отличается от инфицирования клеток животных. Растения имеют прочную клеточную стенку, состоящую из целлюлозы, а грибы — из хитина, так что большинство вирусов могут проникнуть в них только после повреждения клеточной стенки[97]. Однако почти все вирусы растений (включая вирус табачной мозаики) могут перемещаться из клетки в клетку в форме одноцепочечных нуклеопротеиновых комплексов через плазмодесмы[98]. Бактерии, как и растения, имеют крепкую клеточную стенку, которую вирусу, чтобы попасть внутрь, приходится повредить. Но в связи с тем, что клеточная стенка бактерий намного тоньше таковой у растений, некоторые вирусы выработали механизм впрыскивания генома в бактериальную клетку через толщу клеточной стенки, при котором капсид остаётся снаружи[99].
· Лишение оболочек представляет собой процесс потери капсида. Это достигается при помощи вирусных ферментов или ферментов клетки-хозяина, а может быть и результатом простой диссоциации. В конечном счёте вирусная геномная нуклеиновая кислота освобождается.
· Репликация вирусов подразумевает, прежде всего, репликацию генома. Репликация вируса включает синтез мРНК ранних генов вируса (с исключениями для вирусов, содержащих положительную РНК), синтез вирусных белков, возможно, сборку сложных белков и репликацию вирусного генома, которая запускается после активации ранних или регуляторных генов. Вслед за этим может последовать (у комплексных вирусов с крупными геномами) ещё один или несколько кругов дополнительного синтеза мРНК: «поздняя» экспрессия генов приводит к синтезу структурных или вирионных белков.

Отпочковывание вируса от клетки-хозяина
· Вслед за этим происходит сборка вирусных частиц, позже происходят некоторые модификации белков. У вирусов, таких как ВИЧ, такая модификация (иногда называемая созреванием) происходит после выхода вируса из клетки-хозяина[100].
· Выход из клетки. Вирусы могут покинуть клетку после лизиса, процесса, в ходе которого клетка погибает из-за разрыва мембраны и клеточной стенки, если такая есть. Эта особенность есть у многих бактериальных и некоторых животных вирусов. Некоторые вирусы подвергаются лизогенному циклу, где вирусный геном включается путём генетической рекомбинации в специальное место хромосомы клетки-хозяйки. Тогда вирусный геном называется провирусом, или, в случае бактериофага, профагом [101]. Когда клетка делится, вирусный геном также удваивается. В пределах клетки вирус в основном не проявляет себя; однако в некоторый момент провирус или профаг может вызвать активацию вируса, который может вызвать лизис клеток-хозяев[102].
Активно размножающийся вирус не всегда убивает клетку-хозяина. Оболочечные вирусы, в том числе ВИЧ, обычно отделяются от клетки путём отпочковывания. В ходе этого процесса вирус обзаводится своей оболочкой, которая представляет собой модифицированный фрагмент клеточной мембраны хозяина или другой внутренней мембраны[103]. Таким образом, клетка может продолжать жить и продуцировать вирус.